GENETİK
Genetik biyolojinni bir kolu olarak genler, genetik varyasyonu ve kalıtım olarak organizmalar üzerine çalışmalarda bulunan bir bilim dalıdır. [1] [2] [3]
Bin yıl boyunca kalıtım gözlenmiş olmasına rağmen, 19. yüzyılda çalışan bir bilim adamı ve bir keşiş olan Gregor Mendel, genetiği bilimsel olarak inceleyen ilk kişi oldu. Mendel “özellik mirası”, özelliklerin ebeveynlerden yavrulara aktarılma biçiminde kalıplar üzerinde çalıştı. Organizmaların (bezelye bitkileri) ayrık “kalıtım birimleri” yoluyla özellikleri miras aldığını gözlemledi. Bugün hala kullanılan bu terim, gen olarak adlandırılan şeyin bir şekilde belirsiz bir tanımıdır.
Sürekli kalıtım ve moleküler genlerin kalıtım mekanizmaları hala 21. yüzyılda genetiğin birincil ilkeleridir, ama modern genetik fonksiyon ve genlerin davranışlarını inceleyerek için miras ötesinde genişledi. Gen yapısı ve fonksiyonu, varyasyonu ve dağılımı hücre , organizma (örn. Egemenlik ) bağlamında ve bir popülasyon bağlamında incelenir. Genetik, moleküler genetik, epigenetik ve popülasyon genetiği de dahil olmak üzere bir dizi alt alana yol açmıştır. Geniş bir alan içinde incelenmiştir organizmalar yaşam alanları kapsar ( arke , bakteri ve ökaryotlarının).
Genetik süreçler, bir organizmanın çevresi ve gelişime ve davranışa etki eden deneyimlerle birlikte çalışır ve genellikle doğaya karşı yetiştirme olarak adlandırılır . Hücre içi veya hücre dışı bir canlı hücre veya organizmanın çevre veya kapatmak gen transkripsiyonunu geçiş yapabilirler. Klasik bir örnek, biri ılıman bir iklime, diğeri de kurak bir iklime (yeterli şelale veya yağmurdan yoksun) yerleştirilmiş iki genetik olarak özdeş mısır tohumu. İki mısır sapının ortalama yüksekliğinin genetik olarak eşit olduğu belirlenebilirken, kurak iklimdeki iklim, ılıman iklimdeki ortamın su ve besin eksikliğinden dolayı yalnızca yüksekliğinin yarısına kadar büyür.
ETİMOLOJİ
Genetik kelimesi üetken ve oluşum anlamlaı taşıyan antik Yunancada (γενετικός- genetikos) kelimesinden türemiştir. [4] [5] [6]
TARİHÇE
Canlıların ebeveynlerinden özellikleri miras aldığı gözlemi, tarih öncesi dönemlerden beri ekin bitkilerini ve hayvanları seçici üreme yoluyla iyileştirmek için kullanılmıştır . [7] Bu süreci anlamak isteyen modern genetik bilimi, 19. yüzyılda Gregor Mendel’in çalışmasıyla başlamıştır . [8]
Mendel, öncesinde bir Macar soylusu olan Kőszeg “genetik” kelimesini kullanan ilk kişi oldu. Kőszeg, yapıtında genetik kalıtımın çeşitli kurallarını tanımladı. Doğanın genetik yasası (Die genetische Gesätze der Natur, 1819). İkinci yasası Mendel’in yayınladığı kanunla aynıydı. Üçüncü yasasında mutasyonun temel prensiplerini geliştirdi ( Hugo de Vries’in öncüsü olarak kabul edilebilir ). [9]

Harmanlayan miras , mühendis Fleeming Jenkin’in işaret ettiği gibi, doğal seçilim ile evrimi imkansız kılan her bir özelliğin ortalanmasına yol açar .
MENDEL VE KLASİK GENETİK

Morgan’ın Drosophila’da beyaz gözlere neden olan bir mutasyonun cinsiyete bağlı kalıtım gözlemi, onu genlerin kromozomlar üzerinde bulunduğu hipotezine götürdü.
Modern genetik, Mendel’in bitkilerdeki kalıtımın doğası üzerine yaptığı çalışmalarla başladı. Onun kağıt “olarak Pflanzenhybriden über Versuche (” ” Bitki Hibridizasyon üzerinde deneyler için 1865 yılında sunulan”), Naturforschender Verein içinde (Nature Araştırma Derneği) Brunn , Mendel bezelye bitkilerinde bazı özelliklerin kalıtsal modelleri takip ve matematiksel olarak nitelendirdi. [14] Bu kalıtım modeli sadece birkaç özellik için gözlemlenebilse de, Mendel’in çalışması kalıtımın kazanılmış değil partikül olduğunu ve birçok özelliğin kalıtım modellerinin basit kurallar ve oranlarla açıklanabileceğini ileri sürdü.
Mendel’in çalışmasının önemi, ölümünden sonra Hugo de Vries ve diğer bilim adamlarının araştırmasını yeniden keşfettikleri 1900 yılına kadar geniş bir anlayış kazanmadı . Mendel’in çalışmasının bir savunucusu olan William Bateson , 1905’te genetik kelimesini icat etti [15] [16] ( Yunanca genesis —γένεσις, “orijin” kelimesinden türetilen sıfat genetik adından önce geldi ve önce biyolojik anlamda kullanıldı 1860’da [17] ). Bateson hem akıl hocalığı yaptı hem de Cambridge’deki Newnham Koleji’nden diğer bilim adamlarının çalışmaları, özellikle Becky Saunders , Nora Darwin Barlow veMuriel Wheldale Satışta . [18] Bateson kelime kullanımını popüler genetik bitki Hibridizasyon Üçüncü Uluslararası Konferansı yaptığı açılış konuşmasında kalıtım çalışmasını açıklamak için Londra’da 1906 yılında [19]
Mendel’in çalışmasının yeniden keşfinden sonra, bilim adamları hücredeki hangi moleküllerin kalıtımdan sorumlu olduğunu belirlemeye çalıştılar. 1911’de Thomas Hunt Morgan , meyve sineklerindeki cinsiyete bağlı beyaz göz mutasyonunun gözlemlerine dayanarak genlerin kromozomlar üzerinde olduğunu savundu . [20] 1913, onun öğrenci Alfred Sturtevant olgusunu kullanılan genetik bağlantı genleri kromozom doğrusal düzenlenmiştir olduğunu gösterir. [21]
MOLEKÜLER GENETİK

DNA , biyolojik kalıtımın moleküler temeli . Her DNA zinciri, bükülmüş bir merdivende basamaklara benzeyen bir şey oluşturmak için merkezde birbiriyle eşleşen bir nükleotit zinciridir .
James Watson ve Francis Crick , DNA’nın sarmal bir yapıya sahip olduğunu (yani bir tirbuşon şeklinde) gösteren Rosalind Franklin ve Maurice Wilkins’in X-ışını kristalografi çalışmasını kullanarak 1953’te DNA’nın yapısını belirlediler . [25] [26] Çift sarmallı modellerinde iki DNA dizisi vardı ve nükleotidler içe dönüktü, her biri diğer bükümde tamamlayıcı bir nükleotit ile bükülmüş bir merdivende basamaklara benzeyen bir şey oluşturuyordu. [27] Bu yapı genetik bilginin her DNA dizisi üzerindeki nükleotitler dizisinde bulunduğunu gösterdi. Yapı aynı zamanda çoğaltma için basit bir yöntem önerdi: teller ayrılırsa, eski tel dizisine bağlı olarak her biri için yeni partner teller yeniden oluşturulabilir. Bu özellik, DNA’ya, yeni bir DNA ipliğinin orijinal bir ana iplikten olduğu yarı muhafazakar doğasını veren şeydir. [28]
DNA’nın yapısı kalıtımın nasıl çalıştığını göstermesine rağmen, DNA’nın hücrelerin davranışını nasıl etkilediği hala bilinmiyordu. İlerleyen yıllarda, bilim adamları DNA’nın protein üretim sürecini nasıl kontrol ettiğini anlamaya çalıştılar . [29] Hücrenin, uyumlu haberci RNA , DNA’ya çok benzer nükleotidli moleküller oluşturmak için DNA’yı bir şablon olarak kullandığı keşfedildi . Bir haberci RNA’nın nükleotid dizisi, protein içinde bir amino asit dizisi oluşturmak için kullanılır ; nükleotit sekansları ve amino asit sekansları arasındaki bu çeviri genetik kod olarak bilinir . [30]
Yeni ortaya çıkan moleküler kalıtım anlayışı ile bir araştırma patlaması geldi. [31] dikkate değer bir teori kaynaklanan Tomoko Ohta onu değişikliği ile 1973 moleküler evrim nötr teorisi yayın yoluyla moleküler evrim neredeyse nötr teorisi . Bu teoride Ohta, doğal seleksiyonun ve çevrenin önemini genetik evrimin gerçekleşme hızına vurguladı. [32] Önemli bir gelişme, 1977’de Frederick Sanger tarafından zincir sonlandırma DNA dizilimi idi . Bu teknoloji bilim insanlarının bir DNA molekülünün nükleotit sekansını okumasına izin verir. [33] 1983 yılında Kary Banks Mullispolimeraz zincir reaksiyonunu geliştirerek, DNA’nın belirli bir bölümünü bir karışımdan izole etmek ve yükseltmek için hızlı bir yol sağlar. [34] çabaları İnsan Genom Projesi , Enerji, NIH, Bölümü tarafından özel çaba paralel Celera Genomics dizilmesi yol açtı insan genomu 2003 yılında [35] [36]
KATILIM ÖZELLİKLERİ
AYRIK MİRAS VE MENDEL YASALARI

Bir Punnett kare iki mor (B) için heterozigot bezelye bitki ve beyaz (b) çiçekleri çaprazlanmasını gösteren şema
Diploid bir tür olan bezelye durumunda , her bir bitkinin her bir genin iki kopyası vardır, bir kopya her ebeveynden miras alınır. [39] İnsanlar da dahil olmak üzere birçok türün bu kalıtım modeli vardır. Belirli bir genin aynı alelinin iki kopyasına sahip olan diploid organizmalara, bu gen lokusunda homozigot, belirli bir genin iki farklı alleline sahip organizmalara heterozigot denir .
Belirli bir organizma için alel kümesine genotipi, organizmanın gözlemlenebilir özelliklerine ise fenotip denir. Organizmalar bir gende heterozigot olduğunda, nitelikleri organizmanın fenotipine hâkim olduğu için genellikle bir allel baskın olarak adlandırılırken , diğer allel, nitelikleri geri çekildikçe ve gözlemlenmediği için resesif olarak adlandırılır . Bazı allelleri tam hâkim olma ve bunun yerine yok Eksik baskınlık bir ara fenotipi veya kodominantlık anda her iki allelleri şeklinde ifade edilir. [40]
Bir çift organizma cinsel olarak ürediğinde , yavruları her bir ebeveynden iki alelden birini rastgele miras alır. Bu ayrık miras gözlemleri ve alellerin ayrılması, toplu olarak Mendel’in ilk yasası veya Ayrışma Yasası olarak bilinir.
GÖSTERİM VE DİYASGRAM

Genetik soyağacı çizelgeleri, özelliklerin kalıtım kalıplarını izlemeye yardımcı olur.
Genetikçiler kalıtımı tanımlamak için diyagramlar ve semboller kullanırlar. Bir gen, bir veya birkaç harfle temsil edilir. Genellikle bir gen için olağan, mutant olmayan alleli işaretlemek için bir “+” sembolü kullanılır . [41]
Döllenme ve üreme deneylerinde (ve özellikle Mendel’in yasalarını tartışırken) ebeveynlere “P” jenerasyonu ve yavrular “F1” (ilk evlat) nesli olarak adlandırılır. F1 yavruları birbirleri ile çiftleştiğinde, yavrulara “F2” (ikinci evlat) nesli denir. Çapraz üreme sonucunu tahmin etmek için kullanılan yaygın diyagramlardan biri Punnett karesidir .
İnsan genetik hastalıkları üzerinde çalışırken, genetikçiler genellikle özelliklerin mirasını temsil etmek için soyağacı çizelgeleri kullanırlar. [42] Bu çizelgeler, bir soy ağacındaki bir özelliğin mirasını gösterir.
ÇOKLU GEN ETKİLEŞİMLERİ
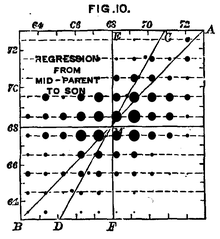
İnsan boyu karmaşık genetik nedenleri olan bir zelliktir. Francis Galton’un 1889’daki verileri, ortalama ebeveyn yüksekliğinin bir fonksiyonu olarak yavru yüksekliği arasındaki ilişkiyi göstermektedir.
Organizmaların binlerce geni vardır ve organizmaların cinsel olarak çoğalmasında bu genler genellikle birbirlerinden bağımsız olarak çeşitlenir. Bu, sarı veya yeşil bezelye rengi için bir alelin kalıtımının, beyaz veya mor çiçekler için alellerin mirasıyla ilgisi olmadığı anlamına gelir. ” Mendel’in ikinci yasası ” veya “bağımsız çeşitlilik yasası ” olarak bilinen bu fenomen, farklı genlerin alellerinin, birçok farklı kombinasyonla yavrular oluşturmak için ebeveynler arasında karıştırıldığı anlamına gelir. (Bazı genler ayırmak, bağımsız bir şekilde, gösteren olmayan genetik bağlantı , bir konu, bu yazıda daha sonra tartışılacaktır.)
Genellikle farklı genler aynı özelliği etkileyecek şekilde etkileşime girebilir. Örneğin Mavi gözlü Mary’de ( Omphalodes verna ), çiçeklerin rengini belirleyen alelleri olan bir gen vardır: mavi veya macenta. Bununla birlikte, başka bir gen, çiçeklerin hiç renge sahip olup olmadığını veya beyaz olup olmadığını kontrol eder. Bir bitkinin bu beyaz alelin iki kopyası olduğunda, çiçekleri beyazdır – ilk genin mavi veya macenta alelleri olup olmadığına bakılmaksızın. Genler arasındaki bu etkileşime epistaz denir , ikinci gen ilke epistatiktir. [43]
Birçok özellik ayrık özellikler değildir (örneğin mor veya beyaz çiçekler), bunun yerine sürekli özelliklerdir (örn. İnsan boyu ve ten rengi ). Bu karmaşık özellikler birçok genin ürünüdür. [44] Bu genlerin etkisi, organizmanın yaşanmıştır ortam tarafından, çeşitli derecelerde, aracılık eder. Bir organizmanın genlerinin karmaşık bir özelliğe katkıda bulunma derecesine kalıtım derecesi denir . [45]Bir özelliğin kalıtsallığının ölçülmesi görecelidir – daha değişken bir ortamda çevrenin, özelliğin toplam varyasyonu üzerinde daha büyük bir etkisi vardır. Örneğin, insan yüksekliği karmaşık nedenlere sahip bir özelliktir. Amerika Birleşik Devletleri’nde% 89 kalıtım derecesi vardır. Bununla birlikte, insanların iyi beslenme ve sağlık hizmetlerine daha değişken bir erişim yaşadığı Nijerya’da, yüksekliğin kalıtılabilirliği sadece% 62’dir. [46]
KALITIM İÇİN MOLEKÜLER TEMEL
DNA VE KROMOZONLAR

Moleküler yapının DNA. Bazlar , teller arasındaki hidrojen bağı düzenlemesiyle eşleşir
DNA normalde çift sarmal şeklinde sarılmış çift sarmallı bir molekül olarak bulunur . DNA’daki her nükleotit, tercihen, karşı iplik üzerinde ortak nükleotid ile eşleşir: T ile bir çift ve G ile C çiftleri. Bu nedenle, iki iplikçik formunda, her iplik, iplikçik şeridi ile birlikte gerekli olan tüm bilgileri etkili bir şekilde içerir. DNA’nın bu yapısı kalıtımın fiziksel temelidir: DNA replikasyonu , telleri bölerek ve her bir ipliği yeni bir partner ipliğinin sentezi için bir şablon olarak kullanarak genetik bilgiyi çoğaltır. [49]
Genler, DNA baz çifti dizilerinin uzun zincirleri boyunca doğrusal olarak düzenlenir. İçinde bakteri , her bir hücre, genellikle tek bir dairesel içeren genophore ise, ökaryotik (örneğin bitkiler ve hayvanlar gibi) organizmaların DNA çoklu doğrusal kromozom düzenlenmiş bulunmaktadır. Bu DNA zincirleri genellikle çok uzundur; örneğin, en büyük insan kromozomu yaklaşık 247 milyon baz çifti uzunluğundadır. [50] Bir kromozomun DNA’sı, DNA’ya erişimi düzenleyen, sıkıştıran ve kontrol eden ve kromatin adı verilen bir materyal oluşturan yapısal proteinlerle ilişkilidir ; ökaryotlarda kromatin genellikle nükleozomlardan , histon çekirdekleri etrafına sarılmış DNA segmentlerinden oluşurproteinler. [51] (bütün kromozom genellikle kombine DNA dizileri), bir organizmadaki kalıtımsal malzemenin bütününe denir genom .
Haploid organizmalar her kromozomun sadece bir kopyasına sahipken , çoğu hayvan ve birçok bitki, her kromozomdan ikisini ve dolayısıyla her genin iki kopyasını içeren diploiddir . [39] Bir gen için iki alel, her biri farklı bir ebeveynden miras alınan iki homolog kromozomun özdeş lokusu üzerinde bulunur .

Walther Flemming’in 1882 ökaryotik hücre bölünmesi diyagramı. Kromozomlar kopyalanır, yoğunlaştırılır ve organize edilir. Daha sonra, hücre bölünürken, kromozom kopyaları kızı hücrelere ayrılır.
ÜREME
Hücreler bölündüğünde, tam genomları kopyalanır ve her yavru hücre bir kopyayı devralır. Mitoz adı verilen bu süreç, en basit üreme şeklidir ve eşeysiz üremenin temelidir . Eşeysiz üreme, çok hücreli organizmalarda da meydana gelebilir ve genomlarını tek bir ebeveynden devralan yavrular üretir. Genetik olarak ebeveynlerine özdeş olan yavrulara klon denir .
Ökaryotik organizmalar genellikle iki farklı ebeveynden miras alınan genetik materyalin bir karışımını içeren yavrular üretmek için cinsel üreme kullanırlar . Cinsel üreme süreci, genomun tek haplarını ( haploid ) ve çift kopyaları ( diploid ) içeren formlar arasında değişir . [39] Haploit hücreler sigorta ve çift kromozomlu diploid hücre oluşturmak için genetik malzemeler. Diploid organizmalar, DNA’larını çoğaltmadan, her bir kromozom çiftinden birini rastgele miras alan kızı hücreleri oluşturmak için bölerek haploidler oluştururlar. Çoğu hayvan ve birçok bitki yaşam sürelerinin çoğu için diploiddir, haploid formu sperm gibi tek hücreli gametlere indirgenmiştir.
Haploid / diploid cinsel üreme yöntemini kullanmasalar da, bakterilerin yeni genetik bilgi edinmenin birçok yöntemi vardır. Bazı bakteriler küçük bir dairesel DNA parçasını başka bir bakteriye aktararak konjugasyona uğrayabilir . [53] Bakteriler, olarak bilinen bir fenomen ortamında bulunan DNA parçalarını alıp genomları içine entegre dönüşümü . [54] Bu süreçler sonucu yatay gen transferi başka alakasız organizmalar arasında genetik bilginin parçalarının nakli. Doğal bakteri dönüşümü birçok bakteriyelde meydana gelir.DNA’yı bir hücreden başka bir hücreye (genellikle aynı türden) aktarmak için cinsel bir süreç olarak kabul edilebilir . [55] dönüştürülmesi çok sayıda bakteriyel etkisini gerektirir gen ürünleri ve özellikle aşağıdaki adaptif bir fonksiyonu olarak belirdiği onarım bölgesinin DNA hasarının alıcı hücrede. [55]
REKOMBİNASYON VE GENETİK BAĞLANTI

Thomas Hunt Morgan’ın 1916 kromozomları arasındaki çift geçişi gösteren resmi.
Geçişin ilk sitolojik gösterimi Harriet Creighton ve Barbara McClintock tarafından 1931’de gerçekleştirildi. Mısır üzerinde yaptıkları araştırma ve deneyler, eşleştirilmiş kromozomlardaki bağlantılı genlerin aslında bir homologdan diğerine değiştiği genetik teori için sitolojik kanıt sağladı. [57]
Kromozomda verilen iki nokta arasında kromozomal geçiş olasılığı, noktalar arasındaki mesafeyle ilişkilidir. Keyfi olarak uzun bir mesafe için, crossover olasılığı genlerin kalıtımının etkili bir şekilde ilişkisiz olacağı kadar yüksektir. [58] yakın genler için, bununla birlikte, genlerin göstermektedir ki geçiş aracının düşük olasılık genetik bağlantı ; iki gen için aleller birlikte miras alınır. Bir dizi gen arasındaki bağlantı miktarları , genlerin kromozom boyunca kabaca düzenlenmesini tanımlayan doğrusal bir bağlantı haritası oluşturmak için birleştirilebilir . [59]
GEN İFADESİ
GENETİK KOD

Genetik kod a kullanarak üçlü kodu DNA, bir geçiş haberci RNA aracı, bir proteini belirtir.
Bu haberci RNA molekülü daha sonra çeviri adı verilen bir işlemle karşılık gelen bir amino asit sekansı üretmek için kullanılır . Kodon adı verilen sekanstaki üç nükleotitin her bir grubu, bir protein içindeki yirmi olası amino asitten birine veya amino asit sekansını sonlandırma talimatına karşılık gelir ; bu yazışmalara genetik kod denir . [60] Bilgi akışı tek yönlüdür: bilgi nükleotit sekanslarından proteinlerin amino asit sekansına aktarılır, ancak asla proteinden tekrar DNA sekansına aktarılmaz – Francis Crick moleküler biyolojinin merkezi dogması olarak adlandırılan bir olgu. [61]
Amino spesifik sekansı asitleri sonuçlar bu protein için eşsiz bir üç boyutlu yapıda ve proteinlerin üç boyutlu yapılar işlevleri ile ilişkilidir. [62] [63] Bazıları protein kolajeninin oluşturduğu lifler gibi basit yapısal moleküllerdir . Proteinler diğer proteinlere ve basit moleküllere bağlanabilir, bazen bağlı moleküller içindeki kimyasal reaksiyonları kolaylaştırarak (proteinin yapısını değiştirmeden) enzim görevi görür . Protein yapısı dinamiktir; protein hemoglobini , memeli kanındaki oksijen moleküllerinin yakalanmasını, taşınmasını ve salınmasını kolaylaştırdığı için biraz farklı formlara dönüşür.
Bir tek nükleotid farkı DNA içinde bir proteinin amino asit sekansı bir değişikliğe neden olabilir. Protein yapıları amino asit sekanslarının bir sonucu olduğu için, bazı değişiklikler yapının stabilitesini bozarak veya proteinin yüzeyini diğer proteinler ve moleküller ile etkileşimini değiştirecek şekilde değiştirerek bir proteinin özelliklerini önemli ölçüde değiştirebilir. Örneğin, orak hücreli anemi , hemoglobinin glo-globin bölümünün kodlama bölgesi içindeki tek bir baz farkından kaynaklanan ve hemoglobinin fiziksel özelliklerini değiştiren tek bir amino asit değişikliğine neden olan bir insan genetik hastalığıdır . [64]Hemoglobinin orak hücre versiyonları , proteini taşıyan kırmızı kan hücrelerinin şeklini bozan lifler oluşturmak için istiflenir . Bu orak şeklindeki hücreler artık kan damarlarından yumuşak bir şekilde akmaz , tıkanma veya parçalanma eğilimi gösterir ve bu hastalıkla ilişkili tıbbi sorunlara neden olur.
Bazı DNA dizileri RNA’ya kopyalanır, ancak protein ürünlerine dönüştürülmez; bu tür RNA moleküllerine kodlayıcı olmayan RNA denir . Bazı durumlarda, bu ürünler kritik hücre fonksiyonlarında (örneğin ribozomal RNA ve transfer RNA ) yer alan yapılara katlanır . RNA ayrıca diğer RNA molekülleri (örn. MicroRNA ) ile hibridizasyon etkileşimleri yoluyla düzenleyici etkilere sahip olabilir .
DOĞA VE BESLENME

Siyam kedileri sıcaklığa duyarlı bir pigment üretim mutasyonuna sahiptir.
Çevre, insan genetik hastalığı fenilketonüri etkilerinde önemli bir rol oynar . [66] Fenilketonüriye neden olan mutasyon, vücudun amino asit fenilalanini parçalama yeteneğini bozar ve ara molekülün toksik bir şekilde birikmesine neden olur ve bu da ilerleyici zihinsel sakatlık ve nöbetlerin ciddi semptomlarına neden olur. Bununla birlikte, fenilketonüri mutasyonuna sahip bir kişi bu amino asidi önleyen katı bir diyet uygularsa, normal ve sağlıklı kalırlar.
Genlerin ve çevrenin (“doğa ve beslenme”) bir fenotipe nasıl katkıda bulunduğunu belirlemek için yaygın bir yöntem, aynı ve kardeş ikizleri veya çoklu doğumların diğer kardeşlerini incelemeyi içerir . [67] aynı zigottan geldiği için Özdeş kardeşler genetik olarak aynıdır. Bu arada, kardeş ikizler normal kardeşler kadar genetik olarak birbirinden farklıdır. Bilim adamları, bir çift özdeş ikizde belirli bir bozukluğun ne sıklıkta meydana geldiğini bir çift çift yumurta ikizinde ne sıklıkta meydana geldiğini karşılaştırarak, bu bozukluğun genetik mi yoksa doğum sonrası çevresel faktörlerden mi kaynaklandığını belirleyebilirler. Ünlü bir örnek , aynı dörtlü olan Genain dörtlülerinin çalışmasını içeriyordu.hepsi şizofreni tanısı aldı . [68] Ancak, bu tür testleri cenin gelişimine etki eden çevresel faktörlerden genetik faktörler ayıramaz.
GEN REGÜLASYONU
Belirli bir organizmanın genomu binlerce gen içerir, ancak bu genlerin herhangi bir anda aktif olmaları gerekmez. Bir gen bir ifade bu mRNA transkripsiyonu ve hücre tarafından gerektiğinde proteinler, sadece üretilir, öyle ki gen ekspresyonunu kontrol birçok hücre yöntemi vardır edilirken. Transkripsiyon faktörleri , bir genin transkripsiyonunu destekleyen veya inhibe eden DNA’ya bağlanan düzenleyici proteinlerdir. [69] genomunda Escherichia coli bakteri, örneğin, amino asit sentezi için gerekli gen bir dizi vardır triptofan. Bununla birlikte, triptofan hücre için zaten mevcut olduğunda, triptofan sentezi için bu genlere artık gerek yoktur. Triptofan varlığı doğrudan genlerin aktivitesini etkiler – triptofan molekülleri triptofan baskılayıcıya (bir transkripsiyon faktörü) bağlanır, baskılayıcının yapısını genin bağlanacağı şekilde değiştirir. Triptofan baskılayıcı, genlerin transkripsiyonunu ve ekspresyonunu bloke eder, böylece triptofan sentez işleminin negatif geri besleme regülasyonunu oluşturur . [70]
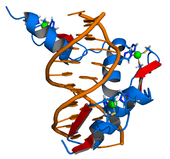
Transkripsiyon faktörleri, ilişkili genlerin transkripsiyonunu etkileyen DNA’ya bağlanır.
Ökaryotlar içinde , genellikle DNA ve kromatinde, kızı hücreler tarafından stabil olarak miras alınan modifikasyonlar şeklinde, genlerin transkripsiyonunu etkileyen kromatinin yapısal özellikleri vardır . [71] Bu özellikler, ” epigenetik ” olarak adlandırılır, çünkü DNA sekansının “üstünde” bulunurlar ve bir hücre oluşumundan diğerine kalıtımı korurlar. Epigenetik özellikler nedeniyle, aynı ortam içinde büyütülen farklı hücre tipleri çok farklı özellikleri koruyabilir. Her ne kadar epigenetik özellikler genellikle gelişim süreci boyunca dinamik olsa da, bazıları, paramutasyon fenomeni gibi, çok nesil kalıtımına sahiptir ve kalıtımın temeli olarak DNA’nın genel kuralında nadir istisnalar olarak bulunurlar. [72]
GENETİK DEĞİŞİM
MUTASYONLAR

Gen çoğalması artıklık sağlayarak çeşitlendirmeye izin verir: bir gen organizmaya zarar vermeden orijinal işlevini değiştirebilir ve kaybedebilir.
DNA replikasyonu işlemi sırasında, zaman zaman ikinci ipliğin polimerizasyonunda hatalar meydana gelir. Mutasyonlar olarak adlandırılan bu hatalar, bir organizmanın fenotipini, özellikle de bir genin protein kodlama dizisi içinde meydana gelirse, etkileyebilir. Hata oranları çok düşüktür – DNA polimerazların “redaksiyon” özelliği nedeniyle her 10-100 milyon bazda bir hata . [73] [74] DNA’daki değişiklik oranını arttıran işlemlere mutajenik denir : mutajenik kimyasallara DNA replikasyonunda, genellikle baz eşleştirme yapısına müdahale ederek hatalar verilirken, UV radyasyonu DNA yapısına zarar vererek mutasyonlara neden olur. . [75]DNA’da kimyasal hasar doğal olarak da meydana gelir ve hücreler uyumsuzlukları ve kopmaları onarmak için DNA onarım mekanizmalarını kullanır. Ancak onarım her zaman orijinal diziyi geri yüklemez. DNA hasarlarının özellikle önemli bir kaynağı hücresel aerobik solunumla üretilen reaktif oksijen türleri [76] gibi görünmektedir ve bunlar mutasyonlara yol açabilir. [77]
DNA değişimi ve genleri yeniden birleştirmek için kromozomal çapraz geçiş kullanan organizmalarda mayoz bölünme sırasındaki hizalama hataları da mutasyonlara neden olabilir. [78] Çaprazlamadaki hatalar, özellikle benzer diziler, ortak kromozomların yanlış bir hizalama benimsemelerine neden olduğunda muhtemeldir; bu, genomlardaki bazı bölgeleri bu şekilde mutasyona daha yatkın hale getirir. Bu hatalar DNA dizisinde büyük değişiklikler yapar – kopyalar , ters çevrmeler , tüm bölgelerin silinmesi – veya farklı kromozomlar arasındaki dizilerin bütün bölümlerinin yanlışlıkla değişimi ( kromozomal translokasyon ).

DOĞAL SELEKSİYON VE EVRİM
Mutasyonlar bir organizmanın genotipini değiştirir ve bu bazen farklı fenotiplerin ortaya çıkmasına neden olur. Çoğu mutasyonun bir organizmanın fenotipi, sağlığı veya üreme zindeliği üzerinde çok az etkisi vardır . [79] Etkisi olan mutasyonlar genellikle zararlıdır, ancak bazen bazıları yararlı olabilir. [80] Sinek Drosophila melanogaster’ deki çalışmalar, bir mutasyon bir gen tarafından üretilen bir proteini değiştirirse, bu mutasyonların yaklaşık yüzde 70’inin geri kalanı nötr veya zayıf olarak faydalı olacak şekilde zararlı olacağını düşündürmektedir. [81]

Bir evrim ağacı ve ökaryotik birkaç karşılaştırılmasıyla inşa organizmalar, ortolog gen dizileri.
Birçok kuşak boyunca, organizmaların genomları önemli ölçüde değişerek evrime yol açabilir . Adaptasyon adı verilen süreçte , yararlı mutasyonlar için seçim, bir türün çevrelerinde daha iyi hayatta kalabilen formlara dönüşmesine neden olabilir. [86] Yeni türler , genellikle nüfusların birbirleriyle gen alışverişi yapmalarını engelleyen coğrafi ayrımlardan kaynaklanan türleşme süreciyle oluşur . [87]
Farklı türlerin genomları arasındaki homolojiyi karşılaştırarak, aralarındaki evrimsel mesafeyi ve ne zaman birbirlerinden ayrıldıklarını hesaplamak mümkündür . Genetik karşılaştırmalar genellikle türler arasındaki ilişkiyi karakterize etmek için fenotipik özelliklerin karşılaştırılmasından daha doğru bir yöntem olarak kabul edilir. Türler arasındaki evrimsel mesafeler, evrimsel ağaçlar oluşturmak için kullanılabilir ; bu ağaçlar , ilişkisiz türler ( yatay gen transferi olarak bilinir ve bakterilerde en yaygın olan) arasında genetik materyalin transferini göstermese de, zaman içinde türlerin ortak inişini ve ayrışmasını temsil eder . [88]
MODEL ORGANİZMALAR

Ortak meyve sineği ( Drosophila melanogaster ) bir popüler model organizma genetik araştırmalarda.
Organizmalar, kısmen rahatlık için seçildi – kısa nesil süreleri ve kolay genetik manipülasyon , bazı organizmaları genetik araştırma araçlarını popüler hale getirdi. Yaygın olarak kullanılan model organizmalar arasında bağırsak bakterisi Escherichia coli , bitki Arabidopsis thaliana , fırıncı mayası ( Saccharomyces cerevisiae ), nematod Caenorhabditis elegans , ortak meyve sineği ( Drosophila melanogaster ) ve ortak ev faresi ( musculus ) bulunur.
TIP
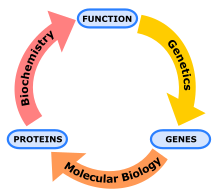
Biyokimya , genetik ve moleküler biyoloji arasındaki şematik ilişki .
Bireyler geliştirmek için kalıtsal eğilim bakımından farklılık kanser , [93] ve kanser genetik bir hastalıktır. [94] Vücuttaki kanser gelişim süreci, olayların bir kombinasyonudur. Mutasyonlar bazen vücuttaki hücreler içinde bölünürken meydana gelir. Bu mutasyonlar herhangi bir yavru tarafından miras alınmayacak olsa da, hücrelerin davranışını etkileyerek bazen daha sık büyümelerine ve bölünmelerine neden olabilirler. Bu süreci durdurmaya çalışan biyolojik mekanizmalar vardır; hücre ölümünü tetiklemesi gereken hücrelere uygunsuz bir şekilde bölünmesi için sinyaller verilir , ancak bazen hücrelerin bu mesajları görmezden gelmesine neden olan ek mutasyonlar meydana gelir. Dahili doğal seçilim sürecivücutta oluşur ve sonunda mutasyonlar, kendi büyümelerini teşvik etmek için hücrelerin içinde birikir ve vücudun çeşitli dokularını büyüten ve istila eden kanserli bir tümör oluşturur .
Normalde, bir hücre sadece büyüme faktörleri olarak adlandırılan sinyallere tepki olarak bölünür ve çevre hücrelerle temas halinde bir kez büyümeyi durdurur ve büyüme engelleyici sinyallere tepki olarak. Genellikle sınırlı sayıda kez bölünür ve ölür, epitel içinde kalarak diğer organlara göç edemez. Bir kanser hücresi olmak için, bir hücrenin birkaç gende (üç ila yedi) mutasyon birikmesi gerekir. Bir kanser hücresi büyüme faktörü olmadan bölünebilir ve inhibitör sinyalleri yoksayar. Ayrıca, ölümsüzdür ve komşu hücrelerle temas ettikten sonra bile süresiz olarak büyüyebilir. Epitelden ve nihayetinde primer tümörden kaçabilir. Daha sonra, kaçan hücre bir kan damarının endotelini geçebilir ve ölümcül metastaz oluşturarak yeni bir organı kolonize etmek için kan dolaşımı yoluyla taşınabilir . Küçük bir kanser fraksiyonunda bazı genetik yatkınlıklar olmasına rağmen, büyük fraksiyon, tümörü oluşturmak için bölünecek ve iletilmeyen bir veya birkaç hücrede orijinal olarak ortaya çıkan ve biriken bir dizi yeni genetik mutasyondan kaynaklanmaktadır. döl ( somatik mutasyonlar ). En sık görülen mutasyonlar, p53 proteini , bir tümör baskılayıcı veya p53 yolunda fonksiyon kaybı ve Ras proteinleri veya diğer onkogenlerde fonksiyon mutasyonlarının kazanılmasıdır .
ARAŞTIRMA YÖNTEMLERİ
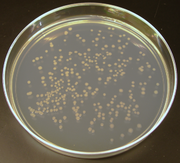
Koloniler, bir E. coli tarafından üretilen hücresel klonlama. Benzer bir metodoloji genellikle moleküler klonlamada kullanılır.
Ligasyon enzimlerinin kullanımı DNA fragmanlarının bağlanmasına izin verir. DNA fragmanlarını farklı kaynaklardan birbirine bağlayarak (“bağlayarak”), araştırmacılar rekombinant DNA oluşturabilirler , DNA genellikle genetik olarak modifiye edilmiş organizmalarla ilişkilendirilir . Rekombinant DNA, plazmidler bağlamında yaygın olarak kullanılır : üzerinde birkaç gen bulunan kısa dairesel DNA molekülleri. Moleküler klonlama olarak bilinen süreçte , araştırmacılar plazmidleri bakterilere ekleyerek ve daha sonra onları agar plakaları üzerinde kültürleyerek (bakteri hücrelerinin klonlarını izole etmek için – “klonlama”, klonlanmış (“klonlama”) çeşitli araçlara başvurabilirler. klonal “) organizmalar).
DNA ayrıca polimeraz zincir reaksiyonu (PCR) adı verilen bir prosedür kullanılarak amplifiye edilebilir . [96] Spesifik kısa DNA dizileri kullanarak PCR, hedeflenen bir DNA bölgesini izole edebilir ve üstel olarak yükseltebilir. Oldukça az miktarda DNA’dan çoğalabildiği için PCR, spesifik DNA sekanslarının varlığını tespit etmek için sıklıkla kullanılır.
DNA DİZİLİMİ VE GENOMİK
Genetiği araştırmak için geliştirilen en temel teknolojilerden biri olan DNA dizilimi , araştırmacıların DNA fragmanlarındaki nükleotit dizisini belirlemelerine olanak tanır. Frederick Sanger tarafından yönetilen bir ekip tarafından 1977’de geliştirilen zincir sonlandırma dizileme tekniği, DNA parçalarını dizmek için rutin olarak kullanılmaktadır. [97] Bu teknolojiyi kullanarak araştırmacılar birçok insan hastalığı ile ilişkili moleküler dizileri inceleyebildiler.
Sekanslama daha ucuz hale geldikçe, araştırmacılar birçok organizmanın genomlarını , birçok farklı fragmandan sekansları birleştirmek için hesaplama araçlarını kullanan genom montajı adı verilen bir işlem kullanarak sekansladılar. [98] Bu teknolojiler dizisine kullanıldı insan genomu içinde İnsan Genom Projesi 2003 yılında tamamlanan [35] Yeni yüksek verim sıralama teknolojileri dramatik birçok araştırmacı a resequencing maliyetini getirmek umuduyla ile DNA dizileme maliyetini düşürüyor insan genomu bin dolara kadar. [99]
Yeni nesil sekanslama (veya yüksek verimli sekanslama), düşük maliyetli sekanslama için sürekli artan talep nedeniyle ortaya çıktı. Bu sekanslama teknolojileri, potansiyel olarak milyonlarca sekansın eşzamanlı olarak üretilmesine izin verir. [100] [101] Mevcut çok sayıda dizi verisi , organizmaların tüm genomlarındaki paternleri araştırmak ve analiz etmek için hesaplama araçlarını kullanan genomik alanını yarattı . Genomik ayrıca büyük biyolojik veri setlerini analiz etmek için hesaplama yaklaşımlarını kullanan bir biyoinformatik alt alanı olarak da düşünülebilir . Bu araştırma alanlarındaki ortak bir sorun, insan özne ile ilgili verilerin nasıl yönetileceği ve paylaşılacağı vekişisel olarak tanımlanabilir bilgiler .
TOPLUM VE KÜLTÜR
19 Mart 2015’te, bir grup önde gelen biyolog , insan genomunu kalıtsal olarak düzenlemek için yöntemlerin, özellikle de CRISPR ve çinko parmağın kullanılmasının dünya çapında yasaklanması yönünde çağrıda bulundu. [102][103][104][105] Nisan 2015’te Çinli araştırmacılar, CRISPR kullanarak yaşayamayan insan embriyolarının DNA’sını düzenlemek için temel araştırma sonuçlarını bildirdiler . [106] [107]
KAYNAKÇA
|
