Kas
| Kas | |
|---|---|

İskelet kasının yukarıdan aşağıya görünümü |
|
| Ayrıntılar | |
| öncü | mesoderm |
| sistem | Kas-iskelet sistemi |
| Tanımlayıcılar | |
| Latin | musculus |
| MeSH | D009132 |
| TA | A04.0.00.000 |
| FMA | 30316 |
Kas , çoğu hayvanda bulunan yumuşak bir dokudur . Kas hücreleri , birbirini geçerek aktin ve miyozinin protein filamanlarını içerir ve hücrenin hem uzunluğunu hem de şeklini değiştiren bir kasılma oluşturur. Kaslar kuvvet ve hareket üretmeye çalışır .
Kas dokuları , miyogenez olarak bilinen bir işlemde embriyonik germ hücrelerinin mezodermal tabakasından türetilir . İskelet veya çizgili, kardiyak ve düz olmak üzere üç tip kas vardır . Kas hareketi gönüllü veya istemsiz olarak sınıflandırılabilir. Kalp ve düz kaslar bilinçli düşünce olmadan kasılır ve istemsiz olarak adlandırılırken, iskelet kasları komuta üzerine kasılır. [1] İskelet kasları da hızlı ve yavaş seğirme liflerine ayrılabilir.
Kaslar ağırlıklı olarak yağların ve karbonhidratların oksidasyonu ile güçlendirilir , ancak özellikle hızlı seğirme lifleri tarafından anaerobik kimyasal reaksiyonlar da kullanılır. Bu kimyasal reaksiyonlar , miyozin kafalarının hareketini güçlendirmek için kullanılan adenosin trifosfat (ATP) moleküllerini üretir . [2]
Kas terimi, belki de bazı kasların şekli veya kasılan kaslar derinin altında hareket eden fareler gibi göründüğü için “küçük fare” anlamına gelen Latin musculus’tan türetilir . [3] [4]
Yapı
Kasların anatomisi , bir organizmanın tüm kaslarını içeren brüt anatomiyi ve tek bir kasın yapılarını içeren mikroanatomiyi içerir.
Türleri
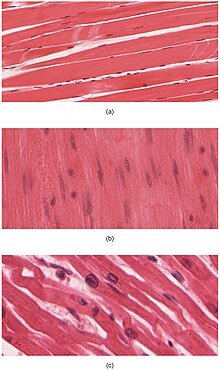
Vücut üç tip kas dokusu içerir: (a) iskelet kası, (b) düz kas ve (c) kalp kası
Kas dokusu, yumuşak doku ve dört temel türlerinden birisidir ve doku, tüm hayvanlarda mevcuttur.
Omurgalı hayvanlarda tanınan üç tip kas dokusu vardır :
- İskelet kası ya da “gönüllü kas” tarafından demirlemiş olan tendon (veya tarafından aponevrozlar birkaç yerlerde) kemik ve efekt için kullanılır iskelet gibi hareketin hareket kabiliyeti ve duruş sürdürülmesinde. Bu postüral kontrol genellikle bilinçsiz bir refleks olarak korunmasına rağmen, sorumlu kaslar postüral olmayan kaslar gibi bilinçli kontrole tepki verir. Ortalama bir yetişkin erkek iskelet kasının% 42’sinden oluşur ve ortalama bir yetişkin kadın% 36’dan oluşur (vücut kütlesinin yüzdesi olarak). [5]
- Düz kas veya “istemsiz kas” yemek borusu, mide, bağırsaklar, bronşlar, rahim, üretra, mesane , kan damarları ve derideki arestör pili (organların ereksiyonunu kontrol ettiği) gibi organların ve yapıların duvarlarında bulunur . vücut kılı). İskelet kasının aksine, düz kas bilinçli kontrol altında değildir.
- Kardiyak kas (miyokard), aynı zamanda bir “istemsiz kas” dır, ancak iskelet kasına yapı bakımından daha benzerdir ve sadece kalpte bulunur.
Kalp ve iskelet kasları, son derece düzenli demetler halinde paketlenmiş sarkomerler içerdikleri için “soyulur” ; düz kas hücrelerinin miyofibrilleri sarkomerlerde düzenlenmez ve dolayısıyla çizgili değildir. İskelet kaslarındaki sarkomerler düzenli, paralel demetler halinde düzenlenirken, kardiyak kas sarkomerler dallanma, düzensiz açılarla (aralanmış diskler olarak adlandırılır) bağlanır. Çizgili kas kasılmaları ve kısa, yoğun patlamalarda gevşemesi, düz kaslar ise daha uzun ve hatta kalıcı kasılmaların devam etmesini sağlar.
İskelet (gönüllü) kas ayrıca iki geniş türe ayrılır : yavaş seğirme ve hızlı seğirme :
- Tip I, yavaş seğirme veya “kırmızı” kas, kılca damarlarla yoğundur ve mitokondri ve miyoglobin açısından zengindir ve kas dokusuna karakteristik kırmızı rengini verir. Yakıt olarak yağ veya karbonhidrat kullanarak daha fazla oksijen taşıyabilir ve aerobik aktiviteyi sürdürebilir . [6] Yavaş seğirme lifleri uzun süre büzülür, ancak çok az kuvvet uygular.
- Tip II, hızlı kas seğirme , üç ana alt tipleri her ikisi de kasılma hızı değişmektedir (İla, IIx ve Ilb) sahiptir [7] oluşturulur ve kuvvet. [6] Hızlı seğirme lifleri hızlı ve güçlü bir şekilde büzülür, ancak çok çabuk yorulur, kas kasılması ağrılı hale gelmeden önce sadece kısa, anaerobik aktivite patlamaları devam eder . En çok kas gücüne katkıda bulunurlar ve kütle artışı için daha fazla potansiyele sahiptirler. Tip IIb, mierokondri ve miyoglobin içinde en az yoğun olan anaerobik, glikolitik , “beyaz” kastır. Küçük hayvanlarda (örn. Kemirgenler) bu, etlerinin soluk rengini açıklayan en hızlı kas tipidir.
Yoğunluk memeli iskelet kas dokusunda 1.06 kg / litre ile ilgilidir. [8] Bu , 0.9196 kg / litre olan yağ dokusu (yağ) yoğunluğuyla kontrastlanabilir . [9] Bu kas dokusunu yağ dokusundan yaklaşık% 15 daha yoğun yapar.
Mikroanatomisi

İskelet kası lifi, kas hücrelerinin sitoplazması olan sarkoplazmayı içeren sarkomimma adı verilen bir plazma membranı ile çevrilidir. Bir kas lifi, hücreye çizgili görünümünü veren birçok fibrilden oluşur.
İskelet kasları , epimysium adı verilen sert bir bağ dokusu tabakası ile kaplanır. Epimysium, kas dokusunu her iki uçta tendonlara tutturur, burada epimysium kalınlaşır ve kollajen olur. Ayrıca kasları diğer kaslara ve kemiklere karşı sürtünmeden korur. Epimysium içinde , her biri topluca bir perimysium tarafından kaplanan 10 ila 100 veya daha fazla kas lifi içeren fasiküller adı verilen birden fazla demet vardır . Her fasikülü çevrelemenin yanı sıra, perimysium sinirler ve kas içindeki kan akışı için bir yoldur. İplik benzeri kas lifleri bireysel kas hücreleridir ( miyositler ) ve her hücre kendi içinde kaplanırendomizyumun bir kolajen lifleri. Böylece, genel kas, kendileri kas oluşturmak için birlikte gruplanan fasiküllere paketlenmiş liflerden (hücreler) oluşur. Her bir demetleme seviyesinde, kollajen bir zar demeti çevreler ve bu zarlar hem dokunun pasif gerilmesine direnerek hem de kaslara uygulanan kuvvetleri dağıtarak kas fonksiyonunu destekler. [10] kas boyunca dağılmış olan kas iğ duyusal geri besleme bilgi merkezi sinir sistemi . (Bu gruplama yapısı epineuryum , perineurium ve endoneurium kullanan sinirlerin organizasyonuna benzer.).
Aynı demetler içinde demetleri yapısı kas hücreleri içinde çoğaltılır . Kas hücreleri içinde, kendileri protein filaman demetleri olan miyofibriller bulunur . “Miyofibril” terimi, bir kas hücresi için başka bir isim olan “myofiber” ile karıştırılmamalıdır. Miyofibriller, sarkomer adı verilen tekrar eden birimler halinde bir araya getirilmiş çeşitli protein filamentlerinden oluşan karmaşık ipliklerdir . Hem iskelet hem de kalp kasının çizgili görünümü, hücrelerindeki normal sarkomer paterninden kaynaklanır. Bu kas türlerinin her ikisi de sarkomer içermesine rağmen, kalp kasındaki lifler tipik olarak bir ağ oluşturmak için dallanır. Kalp kası lifleriaralanmış diskler , [11] bu dokuya bir sinsiyum görünümü verir .
Bir sarkomerdeki filamanlar aktin ve miyozinden oluşur .
Brüt anatomi

Fasikül adı verilen kas lifi demetleri perimysium tarafından kaplıdır. Kas lifleri endomysium ile kaplıdır.
Bir kasın brüt anatomisi, vücuttaki rolünün en önemli göstergesidir. Pennate kaslar ve diğer kaslar arasında önemli bir fark vardır . Çoğu kasta, tüm lifler aynı yöne yönlendirilir, başlangıç noktasından yerleştirmeye kadar bir çizgi halinde ilerler. Bununla birlikte, fıtık kaslarda, tek tek lifler, her iki uçtaki menşe ve yerleştirme tendonlarına yapışan, hareket çizgisine göre bir açıda yönlendirilir. Büzülen lifler kasın genel etkisine bir açıyla çekildiğinden, uzunluktaki değişiklik daha küçüktür, ancak aynı yönelim belirli bir boyuttaki bir kasta daha fazla lif (böylece daha fazla kuvvet) sağlar. Pennate kasları genellikle uzunluk değişikliklerinin rektus femoris gibi maksimum kuvvetten daha az önemli olduğu yerlerde bulunur.
İskelet kası, bir örneği biseps brachii (biseps) olan ayrı kaslarda düzenlenir . İskelet kasının sert, lifli epimizumu tendonlara hem bağlı hem de sürekli . Buna karşılık, tendonlar , kasları çevreleyen periosteum tabakasına bağlanır ve kuvvetin kaslardan iskelete aktarılmasına izin verir. Birlikte, bu lifli tabakalar, tendonlar ve bağlarla birlikte vücudun derin fasyasını oluşturur.
Kas sistemi

Yukarıdaki kas sisteminin ön ve arka görünümlerinde, yüzeysel kaslar (yüzeyde olanlar) vücudun sağ tarafında, derin kaslar (yüzeysel kasların altında olanlar) vücudun sol yarısında gösterilir. Bacaklar için yüzeysel kaslar ön görünümde gösterilirken, arka görünüm hem yüzeysel hem de derin kasları gösterir.
Kas sistemi, tek bir vücutta bulunan tüm kaslardan oluşur. İnsan vücudunda yaklaşık 650 iskelet kası vardır, [12] ancak kesin bir sayının tanımlanması zordur. Zorluk, kısmen farklı kaynakların kasları farklı şekilde gruplandırması ve kısmen de palmaris longus gibi bazı kasların her zaman mevcut olmamasıdır.
Kas kayması , daha büyük bir kas veya kasları arttırmak için hareket eden dar bir kas uzunluğudur.
Kas sistemi, kasları değil, aynı zamanda kemikleri, eklemleri, tendonları ve harekete izin veren diğer yapıları da içeren kas-iskelet sisteminin bir bileşenidir .
Gelişme
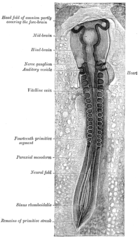
Nöral kıvrımın her iki tarafında paraksiyal mezoderm gösteren bir tavuk embriyosu . Ön (ön) kısım somitler (“ilkel segmentler” olarak etiketlenmiş) oluşturmaya başladı .
Tüm kaslar paraksiyal mezodermden türetilir . Paraksiyal mezoderm, embriyonun uzunluğu boyunca, vücudun segmentasyonuna (en çok vertebral kolonda görülür) karşılık gelen somitlere bölünür . [13] Her somite 3 bölüme sahiptir, sklerotom ( omurları oluşturan ), dermatoma ( cilt) ve miyotom (kas oluşturan) Miyotom , sırasıyla epoksi ve hipaksiyel kasları oluşturan epimere ve hipomere olmak üzere iki bölüme ayrılır İnsanlarda tek epaxial kaslar erector spinalardır.ve küçük intervertebral kaslar ve omurilik sinirlerinin dorsal rami tarafından innerve edilir . Uzuvlar da dahil olmak üzere diğer tüm kaslar hipaksiyaldir ve omurilik sinirlerinin ventral rami tarafından engellenir . [13]
Gelişim sırasında, miyoblastlar (kas progenitör hücreleri) ya omurga ile ilişkili kaslar oluşturmak için somite içinde kalır veya diğer tüm kasları oluşturmak için vücuda göç eder. Miyoblast göçünden önce , genellikle somatik lateral plaka mezoderminden oluşan bağ dokusu çerçeveleri oluşur . Myoblastlar, uzun iskelet kası hücrelerine kaynaştıkları uygun yerlere kimyasal sinyalleri takip eder. [13]
Fizyoloji
Kasılma
Üç kas türü (iskelet, kalp ve düz) önemli farklılıklar gösterir. Ancak, her üç hareketini kullanın aktin karşı miyozin oluşturmak için kasılmasını . İskelet kasında daralma ile uyarılır elektrik darbeleri tarafından iletilen sinirler , motor nöronlar , özellikle (motor sinirler). Kardiyak ve düz kas kasılmaları, düzenli olarak kasılan ve temas ettikleri diğer kas hücrelerine kasılmaları ilerleten dahili kalp pili hücreleri tarafından uyarılır. Tüm iskelet kası ve birçok düz kas kasılması nörotransmitter asetilkolin ile kolaylaştırılır .

Bir sarkomer kasıldığında, Z çizgileri birbirine yaklaşır ve I bandı küçülür. A bandı aynı genişlikte kalır. Tam kasılmada ince ve kalın filamentler üst üste biner
Sinir kontrolü

Temel sinir sistemi fonksiyonunun basitleştirilmiş şeması. Sinyaller duyusal reseptörler tarafından alınır ve periferik sinir sisteminin afferent bacağı yoluyla omuriliğe ve beyne gönderilir, bunun üzerine işlem omuriliğe
Efferent bacak periferal sinir sisteminin kas ve bezlere komutları iletmek için sorumludur ve istemli hareket sonuçta sorumludur. Sinirler , beyinden gelen gönüllü ve otonomik (istemsiz) sinyallere yanıt olarak kasları hareket ettirir . Derin kaslar, yüzeysel kaslar, yüz kasları ve iç kaslar, beynin birincil motor korteksinde , frontal ve parietal lobları bölen merkezi sulkusun hemen önündeki özel bölgelere karşılık gelir .
Ek olarak, kaslar her zaman beyne sinyal göndermeyen refleks sinir sinirlerine tepki verir . Bu durumda, afferent liften gelen sinyal beyne ulaşmaz, ancak omurgadaki efferent sinirlerle doğrudan bağlantılar yoluyla refleksif hareket üretir . Bununla birlikte, kas aktivitesinin çoğunluğu istemlidir ve beynin çeşitli bölgeleri arasındaki karmaşık etkileşimlerin sonucudur.
Memelilerde iskelet kaslarını kontrol eden sinirler , beynin serebral korteksinin primer motor korteksi boyunca nöron gruplarına karşılık gelir . Komutlar da yönlendirilir bazal gangliyonlar ve girişi ile modifiye edilir beyincik üzerinden geçirilmesine önce piramidal yolu ile omurilik ve oradan motor uç plakası kaslarda. Yol boyunca, ekstrapiramidal sisteminki gibi geribildirim, kas tonusunu ve yanıtı etkilemek için sinyallere katkıda bulunur .
Bu tür yer alanlar gibi derin kas duruş genellikle çekirdeklerine kontrol edilir beyin sapı ve bazal gangliyonlar.
Propriyosepsiyon
İskelet kaslarında kas iğleri , kas uzunluğu ve duruş ve eklem pozisyonunun korunmasına yardımcı olmak için merkezi sinir sistemine gerilme hakkında bilgi taşır. Anlamda vücudumuzun uzayda nerede denir propriosepsiyonu , vücut farkındalığı algısı, vücudun çeşitli bölgelerinde herhangi bir anda bulundukları yerleri “bilinçsiz” farkındalık. Beyindeki birçok alan propriosepsiyondan elde edilen geri bildirim bilgileriyle hareketi ve pozisyonu koordine eder. Beyincik ve kırmızı çekirdek özellikle harekete karşı sürekli olarak örnek pozisyon alır ve düzgün hareket sağlamak için küçük düzeltmeler yapar.
Enerji tüketimi
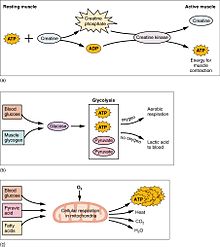
(a) Bazı ATP, dinlenme kasında depolanır. Kasılma başladığında saniyeler içinde tükenir. Kreatin fosfattan yaklaşık 15 saniye daha fazla ATP üretilir. (b) Her glikoz molekülü, aerobik solunumda kullanılabilen veya laktik aside dönüştürülebilen iki ATP ve iki pirüvik asit molekülü üretir.. Oksijen mevcut değilse, piruvik asit, kas yorgunluğuna katkıda bulunabilecek laktik aside dönüştürülür. Bu, yüksek miktarlarda enerjiye ihtiyaç duyulduğu, ancak oksijene yeterince kas verilemediği ağır egzersiz sırasında meydana gelir. (c) Aerobik solunum, karbon dioksit, su ve ATP üretmek için oksijen (O2) varlığında glikozun parçalanmasıdır. Dinlenme veya orta derecede aktif kaslar için gereken ATP’nin yaklaşık yüzde 95’i mitokondride meydana gelen aerobik solunum ile sağlanır.
Kas aktivitesi vücudun enerji tüketiminin çoğunu oluşturur . Tüm kas hücreleri, miyozin kafalarının hareketini güçlendirmek için kullanılan adenosin trifosfat (ATP) molekülleri üretir . Kaslar , ATP’den üretilen ve kreatin kinaz ile gerektiğinde ATP’yi yeniden üretebilen kreatin fosfat formunda kısa süreli bir enerji deposuna sahiptir . Kaslar da şeklinde glikoz depolama form tutmak glikojen . Glikojen hızla glikoza dönüştürülebilirsürdürülebilir, güçlü kasılmalar için enerji gerektiğinde. Gönüllü iskelet kasları içinde glikoz molekülü, işlemde iki ATP ve iki laktik asit molekülü üreten glikoliz adı verilen bir süreçte anaerobik olarak metabolize edilebilir (aerobik koşullarda laktat oluşmadığına dikkat edin; bunun yerine piruvat oluşur ve sitrik asit döngüsü ). Kas hücreleri ayrıca aerobik egzersiz sırasında enerji için kullanılan globüller içerir.. Aerobik enerji sistemlerinin ATP’yi üretmesi ve en yüksek verime ulaşması daha uzun sürer ve daha fazla biyokimyasal adım gerektirir, ancak anaerobik glikolizden çok daha fazla ATP üretir. Diğer yandan kalp kası, üç makro besleyiciden (protein, glikoz ve yağ) herhangi birini ‘ısınma’ süresi olmadan aerobik olarak kolayca tüketebilir ve her zaman dahil olan herhangi bir molekülden maksimum ATP verimini çıkarır. Kalp, karaciğer ve kırmızı kan hücreleri de egzersiz sırasında iskelet kasları tarafından üretilen ve atılan laktik asidi tüketecektir.
İstirahatte iskelet kası günde 54.4 kJ / kg (13.0 kcal / kg) tüketir. Bu, 18.8 kJ / kg (4.5 kcal / kg) ‘da yağ dokusundan (yağ) ve 9.6 kJ / kg (2.3 kcal / kg)’ da kemikten daha büyüktür . [15]
verim
Verim insan kası (bağlamında ölçülmüştür kürek ve bisiklet % 26% 18). Verimlilik, oksijen tüketiminden hesaplanabileceği gibi mekanik iş çıktısının toplam metabolik maliyete oranı olarak tanımlanır . Bu düşük verim elde etme ve yaklaşık 40% verim sonucudur ATP gelen gıda enerjisi, ATP’den enerjinin kas içindeki mekanik çalışmaya dönüştürülmesindeki kayıplar ve vücut içindeki mekanik kayıplar. Son iki kayıp, egzersiz türüne ve kullanılan kas liflerinin türüne (hızlı kasılma veya yavaş kasılma) bağlıdır. Yüzde 20’lik toplam verimlilik için, bir watt mekanik güç saatte 4.3 kcal’a eşdeğerdir. Örneğin, bir kürek ekipmanı üreticisi kürek ergometresini , yakılan kalorileri gerçek mekanik işin dört katına ve saatte 300 kcal’a eşit olacak şekilde kalibre eder , [16]bu, 250 watt mekanik çıkışta yaklaşık yüzde 20 verimlilik demektir. Döngüsel bir kasılmanın mekanik enerji çıkışı, aktivasyon zamanlaması, kas gerginliği yörüngesi ve kuvvet yükselme ve bozulma oranları gibi birçok faktöre bağlı olabilir. Bunlar, çalışma döngüsü analizi kullanılarak deneysel olarak sentezlenebilir .
kuvvet
Kas, üst üste binen üç faktörün bir sonucudur: fizyolojik güç (kas büyüklüğü, kesit alanı, mevcut çapraz köprü, eğitime verilen yanıtlar), nörolojik güç (kasın büzülmesini söyleyen sinyal ne kadar güçlü veya zayıf) ve mekanik güç ( koldaki kas kuvveti açısı, moment kol uzunluğu, eklem yetenekleri). [ alıntı gerekli ]
Fizyolojik güç
| Derece 0 | Kasılma yok |
| 1. derece | Kasılma izi, ancak eklemde hareket yok |
| Derece 2 | Eklem yerçekimi ile hareket ortadan kaldırıldı |
| 3. sınıf | Yer çekimine karşı hareket, ancak ilave dirence karşı değil |
| 4. sınıf | Dış dirence karşı hareket, ancak normalden daha az |
| 5. sınıf | normal güç |
Omurgalı kası tipik olarak izometrik ve optimal uzunlukta olduğunda, kas kesit alanı kare santimetre başına yaklaşık 25–33 N (5,6-7,4 lb f ) kuvvet üretir . [17] yengeç pençelerinde gibi bazı omurgasız kasları, çok daha uzun olan sarkomerlerden çok daha yavaş bir hızda pahasına aktin ve miyozin bağlama ve santimetre kare başına böylece çok daha büyük bir kuvvet için çok daha fazla siteye sonuçlanan omurgalı daha. Bir kasılma tarafından üretilen kuvvet kullanılarak non-invazif olarak ölçülebilir mekanomyografi veya phonomyography , ölçülebilir , in vivo tendon suşu kullanarak (belirgin bir tendon varsa) veya daha invaziv yöntemler kullanılarak doğrudan ölçülebilir.
İskelete uygulanan kuvvet açısından herhangi bir kasın gücü, uzunluğa, kısalma hızına , kesit alanına, parazitlenmeye , sarkomere uzunluğuna, miyozin izoformlarına ve motor ünitelerinin nöral aktivasyonuna bağlıdır . Kas gücünde önemli azalmalar, altta yatan patolojiyi gösterebilir, sağdaki grafik kılavuz olarak kullanılır.
“En güçlü” insan kası
Üç faktör aynı anda kas gücünü etkilediğinden ve kaslar asla bireysel olarak çalışmadığından, bireysel kaslardaki gücü karşılaştırmak yanıltıcıdır ve birinin “en güçlü” olduğunu belirtmek yanıltıcıdır. Ancak aşağıda, farklı nedenlerle gücü dikkat çeken birkaç kas bulunmaktadır.
- Sıradan bir bakış açısına göre, kaslı “güç” genellikle harici bir nesneye bir kuvvet uygulama yeteneğini ifade eder – örneğin, bir ağırlık kaldırma. Bu tanıma göre, masseter veya çene kası en güçlü olanıdır. 1992 Guinness Rekorlar Kitabı, 4 saniyeliğine 4,337 N (975 lb f ) ısırma gücüne ulaştığını kaydediyor . Masseteri ayırt eden şey kasın kendisi hakkında özel bir şey değil, diğer kaslardan daha kısa bir kol koluna karşı çalışmanın avantajıdır.
- “Kuvvet”, kasın kendisi tarafından, örneğin bir kemiğe yerleştirildiği yerde uygulanan kuvveti ifade ediyorsa, en güçlü kaslar en büyük kesit alanına sahip olanlardır. Bunun nedeni, bireysel bir iskelet kası lifinin gerginliğinin çok fazla değişmemesidir. Her lif 0.3 mikronwton civarında bir kuvvet uygulayabilir. Bu tanıma göre, vücudun en güçlü kasının kuadriseps femoris veya gluteus maximus olduğu söylenir .
- Kas kuvveti enine kesit alanı ile belirlendiğinden, daha kısa bir kas , aynı enine kesit alanının daha uzun bir kasından daha fazla “kiloluk pound” (yani ağırlıkça ) olacaktır. Miyometriyal rahim tabaka dişi insan vücudunda ağırlıkça en güçlü kas olabilir. Bir bebeğin doğduğu zamanda , tüm insan rahmi yaklaşık 1,1 kg (40 oz) ağırlığındadır. Doğum sırasında uterus, her kasılma ile 100 ila 400 N (25 ila 100 lbf) aşağı doğru kuvvet uygular.
- Gözün dış kasları, göz küresinin küçük boyutu ve ağırlığı ile ilgili olarak göze çarpan büyüklükte ve güçlüdür . Sıklıkla “yapmak zorunda oldukları iş için en güçlü kaslar” oldukları ve bazen “olması gerekenden 100 kat daha güçlü” oldukları iddia edilir. Bununla birlikte, göz hareketleri (özellikle yüz tarama ve okumada kullanılan sakkadlar ) yüksek hızlı hareketler gerektirir ve hızlı göz hareketi uykusu sırasında göz kasları gece boyunca uygulanır .
- ” Dil vücuttaki en güçlü kastır” ifadesi sıklıkla şaşırtıcı gerçeklerin listelerinde yer alır, ancak bu ifadeyi doğru kılacak herhangi bir “güç” tanımını bulmak zordur. Dilin bir değil sekiz kastan oluştuğunu unutmayın.
- Kalp bir ömür boyunca fiziksel işin büyük miktarda gerçekleştirir kas olmanın bir hak iddia ediyor. İnsan kalbinin güç çıkışının tahminleri 1 ila 5 watt arasındadır . Bu, diğer kasların maksimum güç çıkışından çok daha azdır; örneğin, kuadrisepsler 100 watt’tan fazla üretebilir, ancak sadece birkaç dakika için. Kalp, tüm yaşamı boyunca durmadan aralıksız olarak çalışır ve böylece diğer kasları “dışarı atar”. Seksen yıl boyunca sürekli olarak bir watt çıkışı, iki buçuk gigajoule’lük bir toplam iş çıkışı sağlar . [18]
Egzersiz yapmak
Egzersiz genellikle motor becerileri , zindeliği , kas ve kemik gücünü ve eklem fonksiyonunu geliştirmenin bir yolu olarak önerilir . Egzersizin kaslar, bağ dokusu , kemik ve kasları uyaran sinirler üzerinde çeşitli etkileri vardır . Böyle bir etki, boyutta bir artış olan kas hipertrofisidir . Bu vücut geliştirmede kullanılır .
Çeşitli egzersizler, belirli kas lifi kullanımının bir diğerine göre baskın olmasını gerektirir. Aerobik egzersiz , kasların uzun süre maksimum büzülme mukavemetlerinin çok altında kullanıldığı uzun, düşük efor seviyelerini içerir (en klasik örnek maraton ). Öncelikle aerobik (oksijenli) sisteme dayanan aerobik olaylar, daha yüksek oranda Tip I (veya yavaş seğirme) kas lifleri kullanır, enerji için yağ, protein ve karbonhidrat karışımı tüketir, büyük miktarda oksijen tüketir ve üretir küçük laktik asit. Anaerobik egzersiz , maksimum büzülme mukavemetlerinin çok daha büyük bir yüzdesinde yüksek yoğunluklu kasılmaların kısa patlamalarını içerir. Anaerobik egzersize örnek olarak sprint veağırlık kaldırma . Anaerobik enerji dağıtım sistemi ağırlıklı olarak Tip II veya hızlı seğirme kas lifleri kullanır, esas olarak yakıt için ATP veya glikoza dayanır, nispeten az oksijen, protein ve yağ tüketir, büyük miktarda laktik asit üretir ve uzun bir süre devam edemez aerobik egzersiz olarak. Birçok egzersiz kısmen aerobik ve kısmen anaerobiktir; örneğin, futbol ve kaya tırmanışı her ikisinin bir kombinasyonunu içerir.
Laktik asit varlığı , kas içindeki ATP üretimi üzerinde inhibe edici bir etkiye sahiptir; yorgunluk üretmemesine rağmen, hücre içi konsantrasyon çok yükselirse performansı engelleyebilir ve hatta durdurabilir. Bununla birlikte, uzun süreli eğitim kas içinde neovaskülarizasyona neden olarak atık ürünleri kaslardan çıkarma ve kasılmayı sürdürme yeteneğini arttırır. Sarkomer içinde yüksek konsantrasyonlara sahip kaslardan çıkarıldıktan sonra, laktik asit diğer kaslar veya vücut dokuları tarafından bir enerji kaynağı olarak kullanılabilir veya karaciğere taşınarak pirüvata dönüştürülür.. Laktik asit seviyesini artırmaya ek olarak, yorucu egzersiz kasta potasyum iyonlarının kaybına ve interstisyumda kas liflerine yakın potasyum iyon konsantrasyonlarında bir artışa neden olur. Laktik asit ile asitleme, kuvvetin geri kazanılmasına izin verebilir, böylece asidoz yorgunluğa neden olmaktan ziyade yorgunluğa karşı koruma sağlayabilir. [19]
Gecikmiş kas ağrısı , egzersizden bir ila üç gün sonra hissedilebilecek ağrı veya rahatsızlıktır ve genellikle iki ila üç gün sonra azalır. Bir zamanlar laktik asit birikmesinden kaynaklandığı düşünülürse, daha yeni bir teori, eksantrik kasılma veya alışılmamış eğitim seviyelerinin neden olduğu kas liflerindeki küçük gözyaşlarından kaynaklandığıdır . Laktik asit oldukça hızlı dağıldığından, egzersizden günler sonra yaşanan ağrıyı açıklayamadı. [20]
Klinik önemi
Hipertrofi
Güç ve performans ölçülerinden bağımsız olarak kaslar, hormon sinyali, gelişimsel faktörler, kuvvet antrenmanı ve hastalık gibi bir dizi faktör tarafından daha büyük büyümeye teşvik edilebilir . Popüler inanışın aksine, kas liflerinin sayısı egzersizle artırılamaz . Bunun yerine, kaslar, kas hücresi büyümesinin bir kombinasyonu yoluyla büyür ve mevcut kas hücrelerinin yanında farklılaşmamış uydu hücreleri tarafından sağlanan ilave kütle ile birlikte yeni protein filamentleri eklenir. [12]
Yaş ve hormon seviyeleri gibi biyolojik faktörler kas hipertrofisini etkileyebilir. Boyunca ergenlik erkeklerde, hipertrofi büyüme uyarıcı seviyeleri olarak yüksek bir hızda meydana gelir hormonlar vücut artış üretti. Doğal hipertrofi normalde geç gençlerde tam büyüme ile durur. Gibi testosteron vücudun başlıca büyüme hormonları biridir ortalama olarak erkeklerin kadınlardan daha elde etmek çok daha kolay hipertrofisi bulabilirsiniz. Ek testosteron veya diğer anabolik steroidler almak , kas hipertrofisini artıracaktır.
Kas, spinal ve nöral faktörlerin hepsi kas yapısını etkiler. Bazen bir vücut geliştiricisi sadece sağ pazılara odaklanan bir rejimi tamamladıktan sonra sol pazı daha güçlü bulduğu zaman, bir kişi belirli bir kasta güçte bir artış fark edebilir. Bu fenomene çapraz eğitim denir .
Atrofi

Yetersiz beslenme nedeniyle kas kaybı gösteren savaş esiri. Kaslar yetersiz beslenme, fiziksel hareketsizlik, yaşlanma veya hastalık nedeniyle atrofi olabilir.
İnsanlarda, yatak istirahati veya uzayda uçan astronotlar gibi uzun süreli hareketsiz kalma dönemlerinin kas zayıflaması ve atrofisi ile sonuçlandığı bilinmektedir. Atrofi, insanlı uzay uçuşu topluluğunu özellikle ilgilendirir, çünkü uzay uçuşu sonuçlarında yaşanan ağırlıksızlık, bazı kaslarda kütlenin% 30’u kadar bir kayıptır. [22] [23] Bu gibi sonuçlar altın sarmal zemin sincapları ve kahverengi yarasalar gibi küçük kış uykusuna yatmış memelilerde de görülür. [24]
Yaşlanma sırasında, sarkopeni olarak bilinen iskelet kası fonksiyonunu ve kütlesini sürdürme yeteneğinde kademeli bir azalma vardır . Sarkopeninin kesin nedeni bilinmemektedir, ancak bunun nedeni, “uydu hücrelerinde” iskelet kası liflerinin yenilenmesine yardımcı olan kademeli başarısızlığın ve kritik salgılanan büyüme faktörlerine duyarlılığın azalması veya mevcut olması kas kütlesi ve uydu hücre canlılığını korumak için gereklidir. Sarkopeni, yaşlanmanın normal bir yönüdür ve aslında bir hastalık durumu değildir, ancak yaşlı popülasyondaki birçok yaralanmaya ve yaşam kalitesinin düşmesine neden olabilir. [25]
Kas atrofisine neden olan birçok hastalık ve durum da vardır. Örnekler arasında , kaşeksi adı verilen vücut israfı sendromunu indükleyen kanser ve AIDS sayılabilir . İskelet kası atrofisini indükleyebilen diğer sendromlar veya durumlar konjestif kalp hastalığı ve bazı karaciğer hastalıklarıdır .
Hastalık

Olarak kas distrofisi haline dağınık etkilenen doku ve konsantrasyonu, distrofin (yeşil) büyük ölçüde azalır.
Nöromüsküler hastalıklar kasları ve / veya sinir kontrollerini etkileyen hastalıklardır . Genel olarak, sinir kontrolü ile ilgili problemler, sorunun yeri ve doğasına bağlı olarak spastisiteye veya felce neden olabilir . Büyük bir kısmı, nörolojik bozukluklar arasında değişen, serebrovasküler (felç) ve Parkinson hastalığı için , Creutzfeldt-Jakob hastalığı , hareket ya da sorunlara yol açabilir motor koordinasyon .
Kas hastalıklarının belirtileri zayıflık spastisite, miyoklonus ve miyaljiyi içerebilir . Kas bozukluklarını ortaya çıkarabilecek tanı prosedürleri , kandaki kreatin kinaz seviyelerinin test edilmesini ve elektromiyografiyi (kaslardaki elektriksel aktivitenin ölçülmesi) içerir. Bazı durumlarda, bir miyopatiyi tanımlamak için kas biyopsisi ve ayrıca spesifik miyopatiler ve distrofilerle ilişkili DNA anormalliklerini tanımlamak için genetik test yapılabilir .
Kas gürültüsünü ölçen invaziv olmayan bir elastografi tekniği, nöromüsküler hastalığı izlemenin bir yolunu sağlamak için denemelerden geçmektedir. Bir kas tarafından üretilen ses , kas ekseni boyunca aktomiosin filamanlarının kısalmasından gelir . Kasılma sırasında kas, uzunlamasına ekseni boyunca kısalır ve enine eksen boyunca genişleyerek yüzeyde titreşimler üretir . [26]
Evrim
Metazoanslardaki kas hücrelerinin evrimsel kökeni oldukça tartışılan bir konudur. Bir düşünce dizisinde bilim adamları, kas hücrelerinin bir kez evrimleştiğine ve bu nedenle kas hücreli tüm hayvanların tek bir ortak ataya sahip olduğuna inanıyorlar. Diğer düşünce tarzında, bilim adamları kas hücrelerinin birden fazla evrimleştiğine inanıyor ve herhangi bir morfolojik veya yapısal benzerlik, yakınsak evrimden ve kasın ve hatta mezodermin evriminden önce gelen genlerden kaynaklanıyor – birçok bilim insanının gerçek kas hücrelerine inandığı mikrop tabakası derived.
Schmid ve Seipel, kas hücrelerinin kökeninin, tüm hayvanların sindirim ve sinir sistemlerinin gelişimi ile eşzamanlı olarak meydana gelen monofitik bir özellik olduğunu ve bu kökenin kas hücrelerinin bulunduğu tek bir metazoan atasına kadar takip edilebileceğini savunmaktadır. Bu kaslar, hücreler arası moleküler ve morfolojik benzerlikler olduğunu iddia Cnidaria ve ktenoforun benzer yeterli bilaterians kas hücreleri türetildiği Metazoan’da bir atası olacağını. Bu durumda Schmid ve Seipel, bilateria , ctenophora ve cnidaria’nın son ortak atasının üç mikrop katmanı olan bir triploblast veya bir organizma olduğunu ve bu diploblastinin, iki germ tabakası olan bir organizma anlamına gelir, çoğu cnidarians ve ctenophores’de bulunan mezoderm veya kas eksikliğini gözlemledikleri için ikincil olarak gelişti. Schmid ve Seipel, cnidarians ve ctenophoreslerin morfolojisini bilaterianlarla karşılaştırarak, bazı cnidarians türlerinin ve ctenophores’in dokunaçlarında ve bağırsaklarında miyoblast benzeri yapılar olduğu sonucuna varmışlardır . Bu, kas hücrelerine özgü bir yapı olduğu için, bu bilim adamları akranları tarafından toplanan verilere dayanarak, bunun çift kaslılarda gözlemlenene benzer çizgili kaslar için bir belirleyici olduğunu belirlediler . Yazarlar ayrıca, cnidarians ve ctenophores’de bulunan kas hücrelerinin genellikle bu kas hücrelerinin ektoderm olması nedeniyle yarışmalar olduğunu belirtiyorlar.mezoderm veya mezendoderm yerine. Gerçek kas hücrelerinin kökeni, diğerleri tarafından mezodermin endoderm kısmı olarak tartışılmaktadır.ve endoderm. Bununla birlikte, Schmid ve Seipel, ctenidans’ların medusa aşaması ve polip aşaması ile geliştiğini düşünerek, ktenophores ve cnidarians’ta bulunan kas hücrelerinin gerçek kas hücreleri olup olmadığı konusunda bu şüpheye karşıdır. Hidrozoan medusa aşamasında, çizgili kas hücrelerini mezoderminkine benzer görünecek şekilde oluşturmak ve ektododu bu üçüncü ayrı hücre tabakası olarak adlandırmak için ektodermin distal tarafından ayrılan bir hücre tabakası olduğunu gözlemlerler. . Aynı zamanda, tüm kas hücrelerinin bilaterianlarda mezendodermden türetilmediğini ve anahtar örneklerin hem omurgalıların göz kaslarında hem de spirallerin kaslarında bu hücrelerin endodermal mezodermden ziyade ektodermal mezodermden türediğini iddia ederler. Ayrıca,[27]
Kas hücrelerinin tek bir kökenine yönelik bu argümanın aksine, Steinmetz ve ark. Çizgili kasın bu tek kaynağını belirlemek için kullanılan miyozin II proteini gibi moleküler belirteçlerin aslında kas hücrelerinin oluşumundan önce geldiğini iddia eder . Bu yazar, bu proteini içeren bu çizgili kastan gerçekten yoksun olan porifera veya süngerlerde bulunan kasılma elemanlarının bir örneğini kullanır. Ayrıca, Steinmetz ve ark. bilaterianlarda bulunan ve cnidarians, ctenophores ve bilaterianlarda bulunmayan morfolojik ve moleküler belirteçlerin analizleri yoluyla çizgili kas hücresi gelişiminin polifitik kökenli bir kanıtını sunarlar. Steimetz ve diğ. aktin gibi geleneksel morfolojik ve düzenleyici belirteçlerin, miyozin yan zincirlerinin fosforilasyonunu pozitif kalsiyum konsantrasyonlarının daha yüksek konsantrasyonlarına ve diğer MyHC’ye bağlayabilme yeteneğielementler, sadece kas hücrelerine sahip olduğu gösterilen organizmalarda değil tüm metazoanlarda bulunur. Böylece, bu yapısal veya düzenleyici elemanlardan herhangi birinin, cnidarians ve ctenophores’in kas hücrelerinin, tek bir soyun teyit edilmesi için bilateriyenlerin kas hücrelerine yeterince benzer olup olmadığının belirlenmesinde kullanılması Steinmetz et al. Ayrıca, Steinmetz ve ark. Çizgili kasın kökenini varsaymak için kullanılan MyHc genlerinin ortologlarının, ilk gerçek kas hücrelerini (çizgili kas anlamına gelen) önce gelen bir gen çoğaltma olayı aracılığıyla meydana geldiğini ve MyHc genlerinin süngerlerde bulunduğunu gösterir. kasılma elementleri olan ancak gerçek kas hücreleri olmayan. Ayrıca, Steinmetz ve diğerleri, hem çizgili kas genlerinin oluşumunu kolaylaştırma işlevine hem de hücre düzenleme ve hareket genlerine hizmet eden bu kopyalanmış gen grubunun lokalizasyonunun zaten çizgili myhc ve kas olmayan myhc’ye ayrıldığını gösterdi. Çoğaltılmış gen kümesinin bu ayrılması, çizgili myhc’nin süngerlerde kasılma vakuolüne lokalizasyonu yoluyla gösterilirken, kas olmayan myhc gelişimsel hücre şekli ve değişimi sırasında daha yaygın olarak ifade edildi. Steinmetz ve diğ. cnidarian hariç, cnidarian dışında benzer bir yerelleştirme modeli bulmuşlardır Çoğaltılmış gen kümesinin bu ayrılması, çizgili myhc’nin süngerlerde kasılma vakuolüne lokalizasyonu yoluyla gösterilirken, kas olmayan myhc gelişimsel hücre şekli ve değişimi sırasında daha yaygın olarak ifade edildi. Steinmetz ve diğ. cnidarian hariç, cnidarian dışında benzer bir yerelleştirme modeli bulmuşlardır Çoğaltılmış gen kümesinin bu ayrılması, çizgili myhc’nin süngerlerde kasılma vakuolüne lokalizasyonu yoluyla gösterilirken, kas olmayan myhc gelişimsel hücre şekli ve değişimi sırasında daha yaygın olarak ifade edildi. Steinmetz ve diğ. cnidarian hariç, cnidarian dışında benzer bir yerelleştirme modeli bulmuşlardırSindirim kanalının düz kasında bu çizgili kas markerine sahip N. vectensis . Böylece Steinmetz ve ark. myhc’nin ayrılmış ortologlarının pleiziomorfik özelliğinin kasın monofiyolojisini belirlemek için kullanılamayacağını ve ayrıca bu cnidarianın düz kasında çizgili bir kas markerinin varlığının temel olarak farklı bir kas hücresi gelişimi ve yapısı mekanizması gösterdiğini iddia eder. cnidarians. [28]
Steinmetz ve diğ. bilaterianslarda kas regülasyonu ve oluşumu için troponin kompleksini oluşturmak için kullanılan anahtar bir gen grubunun ve gözlemlenen 47 yapısal ve düzenleyici proteinin eksik olduğunu açıklayarak metazoanslarda çizgili kasın çoklu kökenleri için tartışmaya devam etmek, Steinmetz ve diğ. hem cnidarians hem de bilaterianlarda eksprese edilen benzersiz çizgili kas hücresi proteinini bile bulamadılar. Dahası, Z-diski bilaterianların içinde bile farklı bir şekilde evrimleşmiş gibi görünüyordu ve kas hücreleri için büyük bir radyasyon derecesi gösteren bu klon arasında bile gelişmiş protein çeşitliliği var. Z-diskinin bu ayrışması sayesinde, Steimetz ve diğ. tüm bilaterian kas atalarında mevcut olan sadece dört yaygın protein bileşeninin olduğunu ve bunların gerekli Z-disk bileşenleri için bunların sadece daha önceden tartıştıkları bir aktin proteininin pliziyomorfik durumu aracılığıyla bilgi vermeyen bir belirteç olduğunu savunur. İleri moleküler marker testi ile Steinmetz ve ark. Bilateri olmayanların bilaterians kas oluşumu için gerekli birçok düzenleyici ve yapısal bileşen içermediğini ve süngerler ve amoebozoanlar gibi daha erken, daha ilkel hayvanlarda bulunmayan hem bilaterianlara hem de cnidarians’a ve ctenophores’e benzersiz bir protein seti bulamadıklarını gözlemleyin. Bu analiz sayesinde yazarlar, bilaterians kaslarının yapı ve kullanım için bağımlı olduğu element eksikliğinden dolayı, bilatiyer olmayan kasların farklı bir düzenleyici ve yapısal proteinlerle farklı bir kökene sahip olması gerektiği sonucuna varırlar. [28]
Argümanın bir başka ele alınmasında, Andrikou ve Arnone, gen düzenleyici ağlar üzerindeki yeni mevcut verileri, gen ve morfojenlerin hiyerarşisinin ve diğer doku spesifikasyonu mekanizmasının nasıl ayrıldığını ve erken döterostomlar ve protostomlar arasında nasıl benzer olduğunu araştırmak için kullanır. Tüm bilaterianlarda sadece hangi genlerin mevcut olduğunu değil, aynı zamanda bu genlerin yayılma zamanını ve yerini de anlayarak, Andrikou ve Arnone miyogenezin evriminin daha derin bir anlayışını tartışırlar. [29]
Andrikou ve Arnone makalelerinde kas hücrelerinin evrimini gerçekten anlamak için transkripsiyonel düzenleyicilerin işlevinin diğer dış ve iç etkileşimler bağlamında anlaşılması gerektiğini savunuyorlar. Analizleri sayesinde, Andrikou ve Arnone korunmuş ortologların olduğunu keşfettileromurgasız bilaterianlarda ve cnidarians’ta gen düzenleyici ağın Bu ortak, genel düzenleme devresine sahip olmanın, tek kuyu işleyen bir ağdan yüksek derecede sapmaya izin verdiğini iddia ediyorlar. Andrikou ve Arnone, omurgalılarda bulunan genlerin ortologlarının, omurgasız deuterostom ve protostomlardaki farklı yapısal mutasyon türleri ile değiştirildiğini ve genlerdeki bu yapısal değişikliklerin, kas fonksiyonu ve kas oluşumunda büyük bir farklılaşmaya izin verdiğini iddia etti. bu türler. Andrikou ve Arnone, omurgalılarda ve omurgasızlarda bulunan genlerdeki mutasyon nedeniyle herhangi bir farkı değil, aynı zamanda orijinal gen düzenleyici ağ işlevinden sapmaya neden olabilecek türe özgü genlerin entegrasyonunu da fark edebildi. Böylece, Her ne kadar ortak bir kas modelleme sistemi belirlenmiş olsa da, bunun daha fazla atadan oluşan bir gen düzenleyici ağın, çok farklı kas gelişimine neden olan ilave genler ve mutasyonlar ile soylar boyunca birkaç kez işbirliği yapmasından kaynaklandığını iddia ediyorlar. Dolayısıyla, miyojenik modelleme çerçevesinin atadan kalma bir özellik olabileceği görülmektedir. Bununla birlikte, Andrikou ve Arnone, temel kas modelleme yapısının aynı zamandacis gelişme sırasında farklı zamanlarda bulunan düzenleyici unsurlar . Yüksek seviyede gen ailesi aparatları yapısının aksine, Andrikou ve Arnone, cis düzenleyici elemanların ağdaki zaman ve yerde iyi korunmadığını ve kas hücrelerinin oluşumunda büyük ölçüde farklılık gösterebileceğini bulmuşlardır. Bu analiz sayesinde, miyojenik GRN, miyojenik fonksiyon ve yapıdaki gerçek değişikliklerin, muhtemelen farklı zamanlarda ve yerlerde daha sonraki gen kümeleriyle bağlantılı olan atalardan oluşan bir GRN olduğu görülmektedir. [29]
Evrimsel olarak, iskelet ve kardiyak kasların özel formları omurgalı / eklembacaklı evrim çizgisinin farklılığını ön plana çıkardı . [30] Bu, bu tür kasların 700 milyon yıl önce (mya) bir zaman önce ortak bir atada geliştiğini gösterir . Omurgalı düz kasının iskelet ve kalp kası tiplerinden bağımsız olarak evrimleştiği bulunmuştur.
Referanslar
- ^ Mackenzie, Colin (1918). Kasların Eylemi: Kas Dinlenmesi ve Kasların Yeniden Eğitimi dahil . İngiltere: Paul B. Hoeber. s. 1 . Erişim tarihi: 18 Nisan 2015 .
- ^ Brainard, Jean; Gray-Wilson, Niamh; Harwood, Jessica; Karasov, Corliss; Kraus, Dors; Willan, Jane (2011). CK-12 Ortaokul İçin Yaşam Bilimleri Onurları . CK-12 Vakfı. s. 451 . Erişim tarihi: 18 Nisan 2015 .
- ^ Alfred Carey Carpenter (2007). “Kas” . Anatomi Kelimeler . Erişim tarihi: 3 Ekim 2012 .
- ^ Douglas Harper (2012). “Kas” . Çevrimiçi Etimoloji Sözlüğü . Erişim tarihi: 3 Ekim 2012 .
- ^ Marieb, EN; Hoehn, Katja (2010). İnsan Anatomisi ve Fizyolojisi (8. baskı). San Francisco: Benjamin Cummings. s. 312. ISBN 978-0-8053-9569-3.
- ^ En üste atla:a b McCloud, Aaron (30 Kasım 2011). “Hızlı Seğirme Kas Lifleri Yap” . Komple Vücut Geliştirme . Erişim tarihi: 30 Kasım 2011 .
- ^ Larsson, L; Edström, L; Lindegren, B; Gorza, L; Schiaffino, S (Temmuz 1991). “MHC bileşimi ve yeni bir hızlı seğirme motor ünitesi tipinin enzim-histokimyasal ve fizyolojik özellikleri”. Amerikan Fizyoloji Dergisi . 261 (1 pt 1): C93-101. doi : 10.1152 / ajpcell.1991.261.1.C93 . PMID 1858863 .
- ^ Urbancheka, M; Picken, E; Kalliainen, L; Kuzon, W (2001). “Yaşlı Sıçanların İskelet Kaslarındaki Spesifik Kuvvet Eksikliği Denerve Kas Liflerinin Varlığı ile Kısmen Açıklanmaktadır”. Gerontoloji Serisi A Dergileri: Biyolojik Bilimler ve Tıp Bilimleri . 56 (5): B191-B197. doi : 10.1093 / gerona / 56.5.B191 . PMID 11320099 .
- ^ Farvid, MS; Ng, TW; Chan, DC; Barrett, PH; Watts, GF (2005). “Adiponektin ve resistin yağ dokusu bölmeleri, insülin direnci ve dislipidemi ile ilişkisi”. Diyabet, Obezite ve Metabolizma . 7 (4): 406-413. doi : 10.1111 / j.1463-1326.2004.00410.x . PMID 15955127 .
- ^ MacIntosh, BR; Gardiner, PF; McComas, AJ (2006). “1. Kas Mimarisi ve Kas Elyaf Anatomisi”. İskelet Kası: Biçim ve Fonksiyon (2. bs.). Champaign, IL: İnsan Kinetiği. s. 3-21. ISBN 978-0-7360-4517-9.
- ^ Kent, George C (1987). “11. Kaslar”. Omurgalı Hayvanların Karşılaştırmalı Anatomisi (7. baskı). Dubuque, Iowa: Wm. C. Brown Publishers. s. 326-374 . ISBN 978-0-697-23486-5.
- ^ En üste atla:a b Poole, RM, ed. (1986). İnanılmaz Makine . Washington, DC: National Geographic Topluluğu. sayfa 307-311 . ISBN 978-0-87044-621-4.
- ^ Yukarıçık :a b c Sweeney, Lauren (1997). Embriyolojide Temel Kavramlar: Bir Öğrencinin Hayatta Kalma Rehberi (1. Ciltsiz baskı). McGraw-Hill Profesyonel.
- ^ Kardong, Kenneth (2015). Omurgalılar: Karşılaştırmalı Anatomi, Fonksiyon, Evrim . New York: McGraw Hill Eğitimi. s. 374-377. ISBN 978-1-259-25375-1.
- ^ Heymsfield, SB; Gallagher, D; Kotler, DP; Wang, Z; Allison, DB; Heshka, S (2002). “Dinlenme enerjisi harcamalarının vücut boyutuna bağımlılığı, yağsız kütlenin enerjisel olmayan homojenliğine bağlanabilir”. Amerikan Fizyoloji Dergisi. Endokrinoloji ve Metabolizma . 282 (1): E132 – E138. doi : 10.1152 / ajpendo.2002.282.1.e132 . PMID 11739093 .
- ^ “Concept II Kürek Ergometre, kullanım kılavuzu” (PDF) . 1993 Arşivlenmiş orijinal(PDF) 26 Aralık 2010 tarihinde.
- ^ McGinnis, Peter M. (2013). Spor ve Egzersiz Biyomekaniği (3. baskı). Champaign, IL: İnsan Kinetiği. ISBN 978-0-7360-7966-2.
- ^ Muslumova, Irada (2003). “İnsan Kalbinin Gücü” . Fizik Bilgi Kitabı .
- ^ Nielsen, OB; Paoli, F; Overgaard, K (2001). “Sıçan iskelet kasında laktik asidin kuvvet üretimi üzerindeki koruyucu etkileri” . Fizyoloji Dergisi . 536 (1): 161-166. doi : 10.1111 / j.1469-7793.2001.t01-1-00161.x . PMC 2278832 . PMID 11579166 .
- ^ Robergs, R; Ghiasvand, F; Parker, D (2004). “Egzersize bağlı metabolik asidozun biyokimyası”. J Physiol Regul Integr Comp Physiol kulüpler . 287 (3): R502-516. doi : 10.1152 / ajpregu.00114.2004 . PMID 15308499 .
- ^ Çit, G; Busquets, S; Almendro, V; López-Soriano, FJ; Argilés, JM (2007). “Hibernasyon ayıları plazma plazma antiproteolitik etkileri: kas kaybı tedavisi için yeni bir yaklaşım?”. Clin Nutr . 26 (5): 658-661’de açıklanmaktadır. doi : 10.1016 / j.clnu.2007.07.003 . PMID 17904252 .
- ^ Roy, RR; Baldwin, KM; Edgerton, VR (1996). “Nöromüsküler birimin uzay uçuşuna tepkisi: Sıçan modelinden öğrenilenler”. Exerc. Sport Sci. Rev . 24 : 399-425. doi : 10.1249 / 00003677-199600240-00015 . PMID 8744257 .
- ^ “NASA Kas Atrofisi Araştırması (MARES) Web Sitesi” . Arşivlenmiş orijinal 4 Mayıs 2010 tarihinde.
- ^ Lohuis, TD; Harlow, HJ; Beck, TD (2007). “Hazırda bekletilen siyah ayılar ( Ursus americanus ) kış anoreksi sırasında iskelet kası protein dengesini yaşar”. Zorunlu. Biochem. Physiol. B, Biochem. Mol. Biol . 147 (1): 20-28. doi : 10.1016 / j.cbpb.2006.12.020 . PMID 17307375 .
- ^ Roche, Alex F. (1994). “Sarkopeni: Orta yaşlı ve yaşlılarda ölçümleri ve sağlıkla ilgili önemi üzerine eleştirel bir inceleme”. Amerikan İnsan Biyolojisi Dergisi . 6 (1): 33-42. doi : 10.1002 / ajhb.1310060107 . PMID 28548430 .
- ^ Dumé, Belle (18 Mayıs 2007). ” ‘ Kas gürültüsü’ hastalıkların ilerlemesini ortaya çıkarabilir” . NewScientist.com haber servisi .
- ^ Seipel, Katja; Schmid, Volker (1 Haziran 2005). “Çizgili kasların evrimi: Denizanası ve triploblastinin kökeni”. Gelişimsel Biyoloji . 282 (1): 14-26. doi : 10.1016 / j.ydbio.2005.03.032 . PMID 15936326 .
- ^ Yukarıçık :a b Steinmetz, Patrick RH; Kraus, Johanna EM; Larroux, Claire; Hammel, Jörg U .; Amon-Hassenzahl, Annette; Houliston, Evelyn; Wörheide, Gert; Nikel, Michael; Degnan, Bernard M. (2012). “Cnidarians ve bilaterianlarda çizgili kasların bağımsız gelişimi” . Doğa. 487 (7406): 231-234. doi : 10.1038 / nature11180 . PMC 3398149 . PMID 22763458 .
- ^ Yukarıçık :a b Andrikou, Carmen; Arnone, Maria Ina (1 Mayıs 2015). “Kas yapmanın çok fazla yolu: Miyogenezi yöneten GRN’lerin evrimi”. Zoologischer Anzeiger . Özel Sayı: 3. Uluslararası Omurgasız Morfolojisi Kongresi Bildirileri. 256 : 2-13. doi : 10.1016 / j.jcz.2015.03.005 .
- ^ OOta, S .; Saitou, N. (1999). “Gen ağaçlarının üst üste binmesinden kaynaklanan kas dokularının filogenetik ilişkisi”. Moleküler Biyoloji ve Evrim . 16 (6): 856-867. doi : 10.1093 / oxfordjournals.molbev.a026170 . ISSN 0737-4038 .
