HOMO HEİDELBERGENSİS

Almanya, Heidelberg yakınlarındaki Mauer’den Homo heidelbergenis tipi örneğinin mandibula (alt çene) (Orijinal)
Homo Heilderbergensis veya H. Sapiens Heidelbergensis Orta Pleistosen döneminde ( 300.000 – 600.000 yıl önce) yaşamış olan ve nesli tükenmiş bir insan alt türüdür. 1950’de H. Erectus’un bir alt türü olarak H. Heidelbergensis, ancak yüzyılın sonlarına doğru, kendi türü olarak daha yaygın olarak sınıflandırıldı. H. Heidelbergensis’in yalnızca Avrupa’yla sınırlandırılıp sınırlandırılmayacağı veya Afrika ve Asya örneklerinin de dahile dilip edilmeyeceği tartışılmalıdır ve (Mauer 1) bir çene kemiği olarak karıştırımaktadır çünkü çene kemikleri birkaç tanısal özelliğe sahiptir ve genellikle Orta Pleistosen örnekleri arasında eksiktir. Bundan dolayı örneklerden bazılarının kendi türlerine mi yoksa H. Erectus’un bir alt türüne mi ayrılabileceği tartışılmaktadır. Sınıflandırma çok tartışmalı olduğu için, Orta Pleistosen genellikle “ortadaki karışıklık” olarak adlandırılır.

H. Heidelbergensis yeniden canlandırma
H. Heidelbergensis, Afrika’daki bir H. erectus formundan (bazen H. ergaster olarak da adlandırılır) gelişen bir kronotür olarak kabul edilir. Geleneksel olarak, H. heidelbergensis, modern insanlar (H. sapiens veya H. s. sapiens) ve Neandertaller (H. neanderthalensis veya H. s. neanderthalensis) arasındaki en yakın ortak ata olarak yerleştirilir. H. heidelbergensis’e atanan birçok örnek muhtemelen modern insan/Neandertal ayrımından çok sonra var olmuştur. Orta Pleistosen’de beyin büyüklüğü, modern insanlarla karşılaştırılabilirse ortalama 1.200 cc civarında idi. Orta Pleistosen’deki yükseklik sadece 3 bölgeden kalan kalıntılardan tahmin edilebilir ki Sima de los Huesos, İspanya, erkekler için 169,5 cm ve kadınlar için 157,7 cm Jinniushan, Çin’den bir kadın için 165 cm ve Kabwe, Zambiya’dan bir örnek için 181,2 cm Neandertaller gibi geniş kafatasları vardı ve genel olarak sağlamdılar.
Afrika ve Avrupa’nın Orta Pleistoseni, daha önceki ve çağdaş H. Erectus’tan ayrılan ve muhtemelen artan zeka ile ilişkili olan Geç Aşölyen teknolojisinin ortaya çıkışına işaret eder. Ateş muhtemelen 400.000 yıl önce günlük yaşamın ayrılmaz bir parçası haline geldi ve bu kabaca Avrupa’nın daha kalıcı ve yaygın işgali (45° Kuzey’in üzerinde) ve mızrak oluşturmak için saplama teknolojisinin ortaya çıkışı ile aynı zamana denk geliyor. H. Heidelbergensis koordineli avlanma stratejileri uygulayabilirdi ve benzer şekilde ete daha fazla bağımlı oldukları görülüyor.
TAKSONOMİ
ARAŞTIRMA GEÇMİŞİ
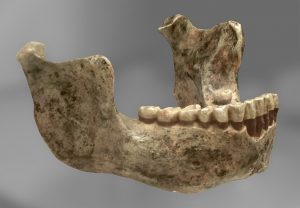
Numune tipi Mauer 1
İlk fosil, Mauer 1 (bir çene kemiği), 1907’de Almanya’nın Heidelberg kentinin güneydoğusunda Mauer’de bir işçi tarafından keşfedildi. Ertesi yıl resmi olarak Alman antropolog Otto Schoetensack tarafından tanımlandı ve onu Homo heidelbergensis adıyla yeni bir tür örneği olarak gösterdi.[1] Bunu, öncelikle mandibulanın arkaikliği (özellikle de muazzam boyutu) nedeniyle yeni bir tür olarak tanımladı ve 640.000 yıllık Avrupa fosil kayıtlarında ki en eski insan çenesiydi. Mandibula iyi korunmuş, sadece sol küçük azı dişleri eksik, 1. sol azı dişinin bir kısmı, sol uçkoronoid işlemi (çene menteşesinde) ve çenenin 2 parça halinde olduğu ve birbirine yapıştırılması gerektiği gibi orta bölümün parçaları ki 3. azı dişindeki hafif aşınmaya göre genç bir yetişkine ait olabilir.[2] 1921’de, Kabwe 1 kafatası İsviçreli madenci Tom Zwiglaar tarafından Kabwe, Zambiya’da (o zamanlar Broken Hill, Kuzey Rhodesia ) keşfedildi ve İngiliz paleontolog Arthur tarafından yeni bir tür olan H. Rhodesiensis olarak tanımlandı.[3]
Bunlar, 20. yüzyılın ilk yarısı boyunca tanımlanan Orta Pleistosen Homo’nun birçok varsayılan türünden ikisiydi. 1950 lerde,Ernst Mayr antropoloji alanına girmişti ve şaşırtıcı bir isim çeşitliliğini araştırarak, yalnızca 3 Homo türü tanımlamaya karar verdi: H. transvaalensis (australopithecines), H. erectus (Mauer çene kemiği ve çeşitli varsayılan Afrika ve Asya taksonları) ve Homo sapiens (modern insanlar ve Neandertaller gibi H. erectus’tan daha genç olanlar dahil). Mayr onları, her türün bir sonra kine (kronotür) evrimleştiği sıralı bir soy olarak tanımladı. Mayr daha sonra australopithecuslar hakkındaki fikrini değiştirmiş olsa da (Australopithecus’u tanıyarak)), onun arkaik insan çeşitliliğine ilişkin daha muhafazakar görüşü, sonraki yıllarda geniş çapta benimsendi.[4]

Rekonstrüksiyonu kafatası Petralona 1 de Ulusal Doğa Tarihi Müzesi
H. erectus hala oldukça değişken, yaygın ve uzun ömürlü bir tür olarak muhafaza edilse de, Orta Pleistosen kalıntılarının tamamının onun içine gömülmesinin haklı olup olmadığı hala çok tartışılıyor. Mayr’ın H.Heidelbergensis’i toplulaştırmasına ilk olarak 1960’da Amerikalı antropolog Francis Clark Howell karşı çıktı. 1974’te İngiliz fiziksel antropolog Chris Stringer, Kabwe 1 ve Yunan Petralona kafatasları arasındaki benzerliklere modern insanların kafataslarıyla (H. sapiens veya H) dikkat çekti. s. sapiens) ve Neandertaller (H. neanderthalensis veya H.s. neanderthalensis). Böylece, Stringer onlarıHomo sapiens sensu lato (geniş anlamda), modern insanların ve Neandertallerin atası olarak. 1979’da Stringer ve Finli antropolog Björn Kurtén, Kabwe ve Petralona kafataslarının Mauer çene kemiği gibi Cromerian endüstrisi ile ilişkili olduğunu bulduve bu nedenle bu üç popülasyonun birbiriyle müttefik olabileceğini öne sürdü. Bu fosiller kötü korunmuş olmalarına ve karşılaştırılabilir pek çok olası tanısal özellik sağlamamalarına rağmen (ve aynı şekilde o zamanlar benzersiz bir türün doğru bir şekilde tanımlanması zordu), en azından bu Orta Pleistosen örneklerinin H. heidelbergensis veya H. rhodesiensis (sırasıyla, Mauer mandibulasının dahil edilmesine veya hariç tutulmasına bağlı olarak) benzerliklerini resmen tanımak zordu.[5]
En etkili şekilde Stringer, paleoantropolog Ian Tattersall ve insan evrimsel biyolog Phillip Rightmire tarafından yapılan daha fazla çalışma, Orta Pleistosen Afro-Avrupa örnekleri ile H. erectus sensu stricto (“katı anlamda”, bu durumda Doğu Asya’dan örnekler) arasında daha fazla farklılık olduğunu bildirdi.[6] Sonuç olarak, 600 ila 300 bin yıl önceki Afro-Avrupa kalıntıları – özellikle Kabwe, Petralona, Bodo ve Arago’dan – genellikle H. heidelbergensis olarak sınıflandırılır. 2010 yılında, Amerikalı fiziksel antropolog Jeffrey H. Schwartz ve Tattersall, tüm Orta Pleistosen Avrupalı ve Asyalı örneklerin sınıflandırılmasını önerdiler.Çin’de Dali ve Jinniushan — H. heidelbergensis olarak.[4] Bu model evrensel olarak kabul görmemiştir. Sibirya’daki bazı benzersiz arkaik insan türlerinin genetik kodunun 2010 yılında tanımlanmasından sonra, teşhis amaçlı fosil buluntuları bekleyen ” Denisovanlar ” olarak adlandırıldı, Asya kalıntılarının aynı türü temsil edebileceği varsayıldı.[5] Böylece, Dali Adamı veya Hint Narmada Adamı gibi Orta Pleistosen Asya örnekleri esrarengiz kalır.[7] Mauer çene kemiğinin 1908’den beri tutulduğu Heidelberg Üniversitesi’ndeki paleontoloji enstitüsü, etiketi H. e. heidelbergensis için2015 yılında H. heidelbergensis.[8]

Avara los huesos de Sima Kafatası 5 (MIGUELON) Natural History Museum, Londra
1976’da İspanya, Sierra de Atapuerca’daki Sima de los Huesos’ta (SH) İspanyol paleontologlar Emiliano Aguirre, José María Basabe ve Trinidad Torres, arkaik insan kalıntılarını kazmaya başladı. Siteyle ilgili araştırmaları, birkaç ayı kalıntısının bulunmasıyla başlatıldı (Ursus deningeri) amatör mağaracılar tarafından 20. yüzyılın başlarından beri (sonuç olarak o bölümdeki bazı insan kalıntılarını yok etti). 1990 yılına kadar yaklaşık 600 insan kalıntısı rapor edildi ve 2004 yılına kadar bu sayı yaklaşık 4.000’e yükseldi. Bunlar, muhtemelen sadece 1’i çocuk olan en az 28 kişiyi ve geri kalanı gençleri ve genç yetişkinleri temsil ediyor. Fosil topluluğu, tüm vücut unsurlarının temsil edildiği tüm cesetlerin hızla gömülmesiyle son derece eksiksizdir.[9] 1997’de İspanyol paleoantropolog Juan Luis Arsuaga bunları H. heidelbergensis’e atadı, ancak 2014’te, Mauer çenesinde bulunan Neandertal benzeri özelliklerin SH insanlarında eksik olduğunu belirterek bunu geri aldı.[10]
SINIFLANDIRMA
Paleoantropolojide, Orta Pleistosen genellikle “ortadaki karışıklık” olarak adlandırılır, çünkü bu dönemden kalma arkaik insan kalıntılarının tür düzeyinde sınıflandırması yoğun bir şekilde tartışılmıştır. Modern insanların (Homo sapiens veya H. s. sapiens) ve Neandertallerin (H. neanderthalensis veya H. s. neanderthalensis) ataları bu zaman diliminde birbirinden ayrıldı ve geleneksel olarak, H. heidelbergensis tipik olarak son ortak ata olarak kabul edilir (LCA). Bu, H. heidelbergensis’i bir kronolojik türün üyesi yapar.[11][7]H. heidelbergensis adının Eski Dünya’daki Orta Pleistosen insanlarını kapsayacak şekilde genişletilip genişletilemeyeceği veya sadece Avrupa ile sınırlandırılmasının daha iyi olup olmadığı çok tartışılıyor. İkinci durumda, Orta Pleistosen Afrika kalıntıları ” H. rhodesiensis ” olarak ayrılabilir.[12][13][14][15] İkinci görüşte, ” H. rhodesiensis ” ya modern insanın doğrudan atası olarak ya da modern insanlara evrimleşen ” H. helmei ” olarak görülebilir.[16]
Orta Pleistosen Avrupa kalıntıları ile ilgili olarak, bazıları Neandertal çizgisine daha sıkı bir şekilde yerleştirilir (yani SH, Pontnewyyd, Steinheim ve Swanscombe), oysa diğerlerinin birkaç benzersiz Neandertal özelliğine sahip olduğu görülüyor (Arago, Ceprano, Vértesszőlős, Bilzingsleben, Mala Balanica ve Aroeira). Bu nedenle, bu bölgede ve zaman diliminde birden fazla soy (veya tür) olduğu öne sürülmüştür, ancak Fransız paleoantropolog Jean-Jacques Hublin bunu haksız bir ekstrapolasyon olarak değerlendirmektedir, çünkü bunlar basitçe farklı olabilir, ancak yine de tek bir yüksek oranda birbirine bağlı popülasyonlar olabilir. değişken türler.[17] 2015 yılında, Marie Antoinette de Lumley, daha az türetilmiş materyalin de kendi türlerine veya H. erectus sl’nin bir alt türüne ayrılabileceğini öne sürdü(örneğin, Arago materyali “H. e. tautavelensis” olarak).[6] 2018’de Mirjana Roksandic ve meslektaşları, H. heidelbergensis’in varsayımını yalnızca Neandertal türevi özelliklere sahip olmayan örnekleri (yani Mauer, Mala Balanica, Ceprano, HaZore’a ve Nadaouiyeh Aïn Askar ) içerecek şekilde revize etti.[18] En son potansiyel H. heidelbergensis arasında tanımlanmış bir ayrım yoktur. malzeme – özellikle Steinheim ve SH – ve en eski Neandertal örnekleri – Biache, Fransa; Ehringsdorf, Almanya; veya Saccopastore, İtalya.[19]
İzole bir çene kemiği olan Mauer mandibulasının, tür için tip örneği olarak kullanılması sorunlu olmuştur, çünkü mandibula birkaç Orta Pleistosen numunesi için eksiktir ve zaten pek çok tanısal özellik sunmuyor. Antropolog William Straus bu konuda “Kafatası Tanrı’nın eseri iken, çene şeytanın eseridir” dedi. Mauer çene kemiği aslında Kabwe kafatasından ve diğer Afro-Avrupa Orta Pleistosen arkaik insanlarından farklı bir türün üyesiyse, o zaman ” H. rhodesiensis ” LCA’nın adı olarak öncelik alacaktır.[7]
EVRİM
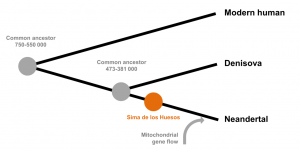
Sima de los Huesos’u kullanan modern insanların ve Neandertallerin türleşmesi devam ediyor.
H. heidelbergensis’in, kabaca 2 milyon yıl önce başlayan homininlerin Afrika dışına ilk erken yayılmaları sırasında – bazen Homo ergaster olarak sınıflandırılan – Afrikalı H. erectus’tan türediği düşünülmektedir. Avrupa çapında dağınık ve Afrika’da Burada kalanlar dönüştü H. Heidelbergensisler veya içine speciated H. Heidelbergensisler Avrupa’da ve ” H. rhodesiensis Afrika’da” ve Doğu Asya’da dağınık olanlar dönüştü H. erectus ss [2] kesin türetme Erken Pleistosen’in sonlarına doğru insan fosil kayıtlarındaki uzun bir boşluk, bir ata türünden türemiştir.[16]2016 yılında Antonio Profico ve meslektaşları, Etiyopya Melka Kunture Formasyonu’nun Gombore II bölgesinden alınan 875.000 yıllık kafatası materyallerinin H. ergaster ve H. heidelbergensis arasında bir geçiş morfunu temsil ettiğini öne sürdüler ve bu nedenle H. heidelbergensis’in Afrika kökenli olduğunu öne sürdüler. Avrupa yerine.[16]
Genetik analize göre, modern insan ve Neandertal’in LCA’sı modern bir insan çizgisine ve bir Neandertal/Denisovan çizgisine ve ikincisi daha sonra Neandertal ve Denisovan’a ayrıldı. Nükleer DNA analizine göre, 430.000 yaşındaki SH insanları, Neandertallerle Denisovalılardan daha yakın akrabadır (ve Neandertal/Denisovan ve dolayısıyla modern insan/Neandertal ayrımının zaten gerçekleşmiş olması), modern insan/Neandertal LCA’nın daha önce meydana geldiğini düşündürür. Arago ve Petralona malzemeleri gibi tipik olarak H. heidelbergensis’e atanan birçok Avrupa örneğinden çok önce vardı.[20]
1997’de İspanyol arkeolog José María Bermúdez de Castro , Arsuaga ve meslektaşları, Gran Dolina, Sierra de Atapuerca’dan gelen kabaca milyon yıllık H. atasını tanımladılar ve LCA için H. heidelbergensis yerine bu türün yerini almayı önerdiler. modern insanlar ve Neandertaller arasında, H. heidelbergensis ondan türemiştir ve yalnızca Neandertallerin atası olan kesinlikle Avrupalı bir türdür.[21] Daha sonra vazgeçtiler.[22] 2020’de Danimarkalı genetikçi Frido Welker ve meslektaşları, bir H. atasından toplanan antik proteinleri analiz ettiler.diş, LCA’nın kendisi olmaktan ziyade LCA’nın kardeş soyunun bir üyesi olduğunu keşfetti (yani, H. heidelbergensis, H. antecessor’dan türememiştir).[23]
45 ° N’nin ötesindeki insan dağılımı, bir milyon yıl önce başlayan kuzeye doğru kısa ömürlü dağılmaların kanıtlarıyla, Alt Paleolitik sırasında oldukça sınırlı görünmektedir. 700.000 yıl önce başlayarak, el baltası teknolojisinin Avrupa’ya yayılmasıyla aynı zamana denk gelen, muhtemelen H. heidelbergensis’in yayılması ve soğuk iklimle başa çıkmak için davranış değişiklikleri ile ilişkili olarak, daha kalıcı popülasyonlar hat boyunca devam etmiş görünüyor. Bu tür işgal, 500.000 yıl öncesinden sonra çok daha sık hale geliyor.[24]
ANATOMİ
KAFATASI

Avara Kabwe 1 at Natural History Museum, Londra
Erken Pleistosen H. erectus/ergaster ile karşılaştırıldığında, Orta Pleistosen insanları çok daha insansı bir yüze sahiptir. Burun açıklığı kafatasında tamamen dikey olarak yerleştirilmiştir ve ön burun eşiği tepeli veya bazen belirgin bir omurga olabilir. Keskin kanallar (üzerinde ağız çatısı dişleri yakınında açık ve) daha yeni insan türünün olduğu gibi yönlendirilir. Frontal kemik geniş, bir parietal kemik genişletilebilir ve temporal kemiğin skuamöz kısmı her artan beyin hacmi ile ilişkili olabilir ki, yüksek ve kemerli olup. sfenoid kemikaşağı doğru uzanan bir omurgaya sahiptir ve kafatasının alt tarafındaki eklem tüberkülü, aksi takdirde çene menteşesinin arkasındaki yüzey oldukça düz olduğu için belirgin bir şekilde dışarı çıkabilir.[25]
2004’te Rightmire, Kabwe, Bodo, Ndutu, Dali, Jinniushan, Petralona, Steinheim, Arago ve SH’den 2 H. heidelbergensis’e atfedilebilecek 10 Orta Pleistosen insanının beyin hacimlerini tahmin etti. Bu set, 1.100 ila 1.390 cc arasında değişen yaklaşık 1.206 cc’lik bir ortalama hacim verir. Ayrıca 30 H. erectus / ergaster beyin hacminin ortalamasını aldı.Doğu Asya ve Afrika’dan yaklaşık 1.5 milyon yıl öncesine yayılan örnekler, 973 cc olarak ve böylece beyin boyutunda önemli bir sıçrama sonuçlandı, ancak kabul edilen beyin boyutu, zamana, coğrafi bölgeye bağlı olarak 727 ila 1.231 cc arasında değişen son derece değişkendi, ve hatta aynı popülasyondaki bireyler arasında (sonuncusu muhtemelen erkeklerden dişilerden çok daha büyük olan belirgin cinsel dimorfizmden dolayı).[25] Karşılaştırıldığında, modern insanlar için ortalama beyin büyüklüğü erkeklerde 1.270 cc ve kadınlarda 1.130 cc’dir; [26] ve Neandertaller için erkekler için 1.600 cc ve kadınlar için 1.300 cc. [27] [28] [29]

Bir büstü H. Heidelbergensisler en Natural History Museum, Londra
2009’da paleontologlar Aurélien Mounier, François Marchal ve Silvana Condemi, Mauer mandibulasını kullanarak H. heidelbergensis’in ilk ayırıcı tanısını ve ayrıca Cezayir, Tighennif’ten gelen materyali yayınladı; SH, İspanya; Arago, Fransa; ve Montmaurin, Fransa. Tanısal özellikleri şu şekilde sıraladılar: küçülmüş bir çene, submental boşlukta (boğaza yakın) bir çentik, yandan görünümde mandibulanın paralel üst ve alt sınırları, yanak dişlerinin yakınında birkaç mental foramen (kan damarları için küçük delikler), yatay bir retromolar boşluk (azı dişlerinin arkasında bir boşluk), azı dişleri ve ramus (kafatasına bağlanmak için yukarı çıkan) arasında bir oluk ), genel bir uzun çene, derinMasseter kası (çeneyi kapatan) için fossa (bir çöküntü), küçük bir gonial açı (mandibula gövdesi ile ramus arasındaki açı), geniş bir planum alveolare (en öndeki diş yuvasından arkaya olan mesafe) çene), gelişmiş bir planum üçgen (çene menteşesine yakın) ve 3. molar seviyesinden çıkan bir milohyoid çizgi. [2]
Oluştur
Orta Pleistosen boyunca vücut büyüklüğündeki eğilimler, genel olarak uzuv kemikleri ve kafatası olmayan (kafatası sonrası) kalıntılardan dolayı belirsizdir. Çeşitli uzun kemiklerin uzunluklarına dayalı olarak, SH insanları erkeklerde ortalama kabaca 169,5 cm ve dişiler için 157,7 cm, maksimum sırasıyla 177 cm ve 160 cm. Jinniushan’dan bir dişi kısmi iskeletin yüksekliğinin, yaşamda yaklaşık 165 cmolarak oldukça uzun olduğu ve SH dişilerinden çok daha uzun olduğu tahmin edilmektedir. Kabwe’den bir kaval kemiğinin tipik olarak en uzun Orta Pleistosen örnekleri arasında 181,2 cm olduğu tahmin edilmektedir, ancak bu bireyin ya alışılmadık derecede büyük olması ya da femurdan çok daha uzun bir kaval kemiğine sahip olması mümkündür.oran beklenenden daha fazla. Bu örnekler kendi kıtalarını temsil ediyorsa, Orta Pleistosen Eski Dünyası boyunca orta ila uzun boylu insanların yaygın olduğunu öne sürerler. Durum buysa, o zaman herhangi bir arkaik insan türünün tüm popülasyonlarının ortalama yüksekliği genellikle 165-170 cm olacaktır. Skhul ve Qafzeh kalıntıları erkekler için ortalama 185.1 cm ve kadınlar için 169.8 cm, ortalama 177.5 cm ile erken modern insanlar özellikle daha uzundu. daha uzun bacaklarla uzun mesafeli seyahatin enerji verimliliğini artırın.[30] Namibya’daki Berg Aukas Madeni’nden yaklaşık 20 km doğusunda bulunan belirgin şekilde masif bir proksimal (üst yarı) femur bulundu. Grootfontein’in başlangıçta 93 kg kadar olduğu tahmin ediliyordu, ancak fahiş boyut şimdi olgunlaşırken yoğun aktivite seviyesine bağlanıyor; Berg Aukas bireyi muhtemelen orantısal olarak Kabwe 1’e benziyordu.[31]
İnsan bauplanı (vücut planı) H. ergaster’de evrimleşmiştir ve daha sonraki tüm Homo türlerini karakterize eder, ancak daha türetilmiş üyeler arasında 2 farklı morf vardır: modern insanlar gibi dar göğüslü ve zarif bir yapı ve daha geniş göğüslü ve Neandertaller gibi sağlam yapı. Bir zamanlar, ince yapılı H. ergaster kısmi iskeleti KNM WT-15000’e (Turkana Boy dayanarak Neandertal yapısının Neandertallere özgü olduğu varsayılmıştı.), ancak bazı Orta Pleistosen iskelet unsurlarının keşfi (genel olarak parçalı ve uzak ve az sayıda olsa da) Orta Pleistosen insanlarının genel olarak daha fazla Neandertal morfu içerdiğini gösteriyor. Bu, son derece iyi korunmuş SH topluluğunda en açık şekilde gösterilmiştir.Kafatasının sağlamlığına dayanarak, Orta Pleistosen insanlarının yüksek derecede cinsel dimorfizme sahip olduğu varsayılmıştır, ancak SH insanları modern insan benzeri bir seviye göstermektedir.[32]
SH insanları ve diğer Orta Pleistosen Homo, daha fazla bazal pelvise ve femurlara sahiptir (Neandertallerden daha erken Homo’ya benzer). Genel olarak geniş ve eliptik pelvis, Neandertallerin veya modern insanlarınkinden daha geniş, daha uzun ve daha kalındır (ön-arkaya doğru genişlemiştir) ve öne yerleştirilmiş bir asetabulokristal payandaya (kalça kaçırma sırasında iliak krestleri destekler), iyi tanımlanmış bir supraasetabular oluğa (arasında) sahiptir. kalça soketi ve ilium) ve ince ve dikdörtgen bir superior kasık ramusu (modern insanlarda kalın, sağlam olanın aksine). Tüm arkaik insanların ayaklarında ayak bileği kemiğinden daha uzun bir troklea bulunur., ayak bileğini daha esnek hale getirir (özellikle dorsifleksiyon ve plantar fleksiyon).[32]
PATOLOJİ
Yüzünün sol tarafında, bir SH kafatası (Kafatası 5) bilinen en eski orbital selülit vakasını (ağızdaki bir apseden gelişen göz enfeksiyonu) sunar. Bu muhtemelen sepsise neden oldu ve kişiyi öldürdü.[33] [34] [35]
Eklem dejenerasyonuna dayanan bir erkek SH pelvisi (Pelvis 1) 45 yıldan fazla yaşamış olabilir ve bu da onu insan fosil kayıtlarındaki bu demografinin en eski örneklerinden biri yapar. 45+ bireyin sıklığı zamanla kademeli olarak artar, ancak genel olarak Paleolitik boyunca oldukça düşük kalmıştır. Benzer şekilde yaşa bağlı hastalıklar lomber kifoz (beldeki lomber vertebranın aşırı eğriliği), L5-S1 spondilolistezis (son lomber vertebranın ilk sakral vertebra ile yanlış hizalanması ) ve L4 ve 5’te Baastrup hastalığı (genişleme) vardı. spinöz süreçler). Bunlar, hareketi önemli ölçüde sınırlayan bel ağrısına neden olabilir ve grup bakımının kanıtı olabilir.[36]
Bir ergen SH kafatasına (Cranium 14), lambdoid tek sütürlü kraniyosinostoz teşhisi kondu (sol lambdoid sütürünün olgunlaşmamış kapanması, gelişme devam ederken kafatası deformitelerine yol açar). Bu, modern insanlarda her 200.000 kişiden 6’sından azında meydana gelen nadir bir durumdur. Birey 5 yaşına yakın bir zamanda öldü, bu da deformasyonu nedeniyle tarihsel zamanlarda yapıldığı gibi terk edilmediğini ve diğer çocuklarla aynı kalitede bakım aldığını gösteriyor.[37]
Dişlerdeki mine hipoplazisi, beslenme stresi nöbetlerini belirlemek için kullanılır. SH insanlar için% 40 oranda, bu daha önceki Güney Afrika sergilenen daha yüksektir insanımsı Paranthropus robustus de Swartkrans (% 30.6) ya da Sterkfontein (12.1%). Bununla birlikte, Neandertaller daha da yüksek oranlara ve daha yoğun hipoplazi nöbetlerine maruz kaldılar, ancak bunun Neandertallerin doğal kaynakları daha az kullanma yeteneklerinden mi yoksa daha zorlu ortamlarda yaşamalarından mı kaynaklandığı açık değil. 3.5 yaşında bir zirve, sütten kesme yaşı ile ilişkili olabilir. Neandertallerde bu zirve 4 yaşındaydı ve birçok modern avcı toplayıcı da yaklaşık 4 yaşında sütten kesiliyor.[38]
KÜLTÜR
GIDA

Amédée Forestier tarafından ” H. rhodesiensis ” kabilesinin 1922 restorasyonu
Genel olarak Orta Pleistosen toplulukları, büyük bir oyunu öncekilerden daha yüksek bir sıklıkta yemiş gibi görünüyor ve et, temel bir diyet bileşeni haline geldi. Örneğin Terra Amata sakinleri ağırlıklı olarak geyik yeme gibi görünüyor, ama aynı zamanda filler, yaban domuzu, yaban keçisi, gergedan ve – Diyet genel farklı şekillerde olabilir yaban öküzü. Afrika bölgeleri tipik olarak yaygın olarak sığır ve at kemikleri verir. Karkaslar basitçe temizlenmiş olsa da, bazı Afro-Avrupa bölgeleri tek bir türün spesifik olarak hedeflendiğini gösteriyor ve bu da daha çok aktif avlanmayı gösteriyor; örneğin: 50 ila 60’tan fazla bireysel babun veren Olorgesailie, Kenya (Theropithecus oswaldi); ve İspanya’da bol miktarda fil kemiğine sahip Torralba ve Ambrona (gergedan ve büyük toynaklı memeliler de dahil). Et geçimindeki artış, Orta Pleistosen’de grup avcılığı stratejilerinin gelişimini gösterebilir. Örneğin, Torralba ve Ambrona’da hayvanlar öldürülmeden önce bataklıklara koşmuş olabilir, bu da büyük bir avcı grubu tarafından koordineli ve organize bir saldırıyla kuşatılıp sürülmesini gerektirmiştir. Bazı sahaların okyanusa, göllere veya nehirlere yakın olmasına rağmen, su ortamlarının işletilmesi genellikle oldukça eksiktir.[39]
Bitkiler muhtemelen mevsimsel olarak temin edilebilenler de dahil olmak üzere sıklıkla tüketiliyordu, ancak hayvan kemikleri kadar fosilleşmedikleri için sömürülmelerinin boyutu belirsizdir. Yağsız et ağırlıklı bir diyet varsayarsak, bir bireyin protein zehirlenmesini önlemek için, tipik olarak bol miktarda yeraltı depolama organları, ağaç kabuğu, çilek veya fındık yiyerek yüksek karbonhidrat alımına ihtiyacı olacaktır. Schöningen sitesi, Almanya, ya çiğ yenilebilir veya pişmiş zaman vardır çevresinde 200’ün üzerinde tesisi bulunmaktadır.[40]
SANAT
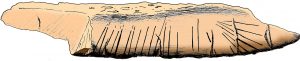
Almanya, Bilzingsleben’den 370.000 yıllık oyulmuş fil kaval kemiğinin çizimi
Üst Paleolitik çağın modern insanları, sembolik değeri olan kazınmış gravürlere sahip oldukları için iyi bilinirler. 2018 itibariyle, yalnızca 27 Orta ve Alt Paleolitik nesnenin sembolik gravürlere sahip olduğu varsayılmıştır ve bunlardan bazılarının doğal veya sembolik olmayan olaylardan (fosilleşme veya kazı süreçleri gibi) kaynaklandığı reddedilmiştir. Alt Paleolitik olanlar şunlardır: Terra Amata’dan 380.000 yıllık üç çakıl; Almanya, Markkleeberg’den 250.000 yıllık bir çakıl taşı; Lazaret’ten yaklaşık 200.000 yıllık 18 çakıl taşı (Terra Amata yakınlarında); Monako’da ki Grotte de l’Observatoire’dan yaklaşık 200.000 yıllık bir litik; Bilzingsleben’den 370.000 yıllık bir kemik, Almanya ve Baume Bonne, Fransa’dan 200 ila 130 bin yıllık bir çakıl taşı.[41]
19. yüzyılın ortalarında, Fransız arkeolog Jacques Boucher de Crèvecœur de Perthes, Fransa’nın Amiens kentindeki St. Acheul’da (Acheulian’ın tanımlandığı alan) kazıya başladı ve el baltalarına ek olarak delikli sünger fosilleri bildirdi (Porosphaera). globularis) dekoratif boncuklar olduğunu düşündü. Bu iddia tamamen göz ardı edildi. 1894’te İngiliz arkeolog Worthington George Smith, Bedfordshire’da benzer 200 delikli fosil keşfetti., İngiltere ve ayrıca Boucher de Perthes’in bulgusuna atıfta bulunmasa da, muhtemelen bunun farkında olmadığı için işlevlerinin boncuk olduğunu tahmin etti. 2005 yılında, Robert Bednarik malzemeyi yeniden inceledi ve – tüm Bedfordshire P. globularis fosilleri alt küresel olduğundan ve bu türün oldukça değişken bir şekle sahip olmasına rağmen, çapları 10-18 mm arasında değiştiği için – şu sonuca vardı: bilinçli olarak seçilmişlerdir. Bazı parazitik yaratıklar tarafından (yani doğal süreçler yoluyla) tamamen veya neredeyse tamamen sıkılmış gibi görünüyorlar ve daha sonra deliği tamamen açmak için daha kapalı olan ucun ne olacağı konusunda vuruldular. Ayrıca, diğer boncukların birbirine bağlanıp bir kolye olarak takıldığında takırdamaktan doğduğunu tahmin ettiği aşınma fasetleri de buldu.[42] 2009 yılında, Solange Rigaud, Francisco d’Errico ve meslektaşları, değiştirilmiş alanların, değiştirilmemiş alanlara göre daha açık renkli olduğunu fark ettiler ve bu, kazı sırasında olduğu gibi çok daha yakın bir zamanda uygulandıklarını düşündürdü. Ayrıca, fosillerin, bölgelerdeki Acheulian eserleriyle güvenle ilişkilendirilebileceğine ikna olmadılar ve – arkaik insan faaliyetlerine bir alternatif olarak – görünen boyut seçiminin, ya doğal jeolojik süreçlerden ya da bunu tercih eden 19. yüzyıl koleksiyoncularından kaynaklanabileceğini öne sürdüler. özel formu. [43]
Erken modern insanlar ve geç Neandertaller (ikincisi özellikle 60.000 yıl öncesinden sonra), kana benzer bir renk ürettiği için muhtemelen sembolik amaçlar için kırmızı aşı boyasını yaygın olarak kullandılar, ancak aşı boyasının işlevsel bir tıbbi uygulaması da olabilir. Bu iki türün ötesinde, iki kırmızı hardal topaklarının bulunduğu Olduvai Gorge, Tanzanya’da hardal rengi kullanımı kaydedilmiştir; Bir hardal rengi levhanın belirli bir şekle budandığı Ambrona; ve sarıdan kırmızı-kahverengiye ve kırmızıya kadar geniş bir renk aralığı elde etmek için 75 hardal rengi parçanın ısıtıldığı Terra Amata. Bunlar, erken ve izole renk tercihi ve renk sınıflandırması örnekleri olabilir ve bu tür uygulamalar henüz normalleştirilmemiş olabilir.[44]
2006’da Eudald Carbonell ve Marina Mosquera, SH homininlerinin mağaraya girme gibi bazı feci bir olayın kurbanı olmak yerine insanlar tarafından gömüldüğünü öne sürdüler, çünkü küçük çocuklar ve bebekler ortada yok, bu tek ve eksiksiz olsaydı beklenmedik olurdu. aile birimi. SH insanları dikkat çekici bir şekilde yalnızca tek bir taş aletle, yüksek kaliteli kuvarsitten (bölgede nadiren kullanılan) yapılmış özenle hazırlanmış bir el baltasıyla ilişkilendirilir ve bu nedenle Carbonell ve Mosquera, bunun bedenlerle kasıtlı ve sembolik olarak bazı nesneler olarak yerleştirildiğini öne sürdüler. tür mezar iyi. Sembolik mezarların sözde kanıtları 300.000 yıl daha su yüzüne çıkmayacaktı. [45]
TEKNOLOJİ
TAŞ ALETLER
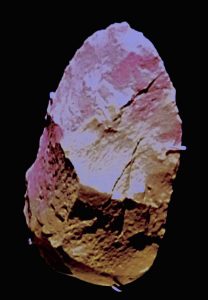
Sima de los huesos elle axe
uzun 15.5 sm (6 inç) [45]
Alt Paleolitik (Erken taş devri) içermesidir Oldowan ile değiştirildi Acheulian çok simetrik üretimi ile karakterize elle eksenleri. Acheulian’ın yaklaşık bir milyon yıllık bir zaman aralığı vardır ve bu tür teknolojik durgunluk, tipik olarak, bilişsel akışkanlıktaki eksiklik, çalışma belleği gibi yenilikçi kapasiteyi önemli ölçüde azaltan nispeten sınırlı bilişsel yeteneklere atfedilmiştir.veya çıraklıkla uyumlu bir sosyal sistem. Bununla birlikte, Acheulian zaman içinde ustaca değişiyor gibi görünüyor ve tipik olarak Erken Acheulian ve Geç Acheulian olarak ikiye ayrılıyor, ikincisi özellikle 600 ila 500 bin yıl önce popüler hale geliyor. Geç Acheulian teknolojisi, Movius Hattı’nın doğusundan Doğu Asya’ya asla geçmedi, bunun genellikle kültürel aktarımdaki bazı büyük eksikliklerden (yani Doğu’daki daha küçük nüfus büyüklüğü)[46] ya da çok daha az taş kadar koruma önyargısından kaynaklandığına inanılıyor. alet toplulukları hattın doğusunda bulunur. [47]
Geçiş, daha küçük, daha ince ve daha simetrik el eksenlerinin üretimi ile gösterilir (ancak daha kalın, daha az rafine olanlar hala üretildi). İngiltere’deki 500.000 yıllık Boxgrove sahasında – çok sayıda alet kalıntısı bulunan son derece iyi korunmuş bir bölge – el baltasına yumuşak bir çekiçle neredeyse dik olarak vurularak inceltme yapılmış olabilir, alet yapımı için hazırlanmış platformların icadıyla mümkün olabilir. Boxgrove knappers, muhtemelen daha sonra başka aletlere geri dönüştürmek amacıyla, el baltalarının yapımından arta kalan büyük litik pulları da geride bıraktı. Geç Acheulian siteleri başka yerlerde önceden hazırlanmış litik çekirdekler (“Large Flake Blanks,” LFB) çeşitli şekillerde araçlara dönüştürmeden önce, hazırlanmış platformları gereksiz hale getirir. LFB Acheulian, bir milyon yıl önce Afrika’dan Batı ve Güney Asya’ya yayılmıştır ve 600.000 yıl önce Güney Avrupa’da bulunur, ancak kuzey Avrupa (ve 700.000 yıl önce Levant ) çoğunlukla küçük, kalın çakmaktaşı nodüller. Afrika’da hazırlanan ilk platformlar, Erken Taş Devri (Acheulian) ile Orta Taş Devri arasında geçiş yapan 450.000 yıllık Fauresmith endüstrisinden gelmektedir.[46]

Boxgrove, İngiltere’de bulunan yüzlerce el baltasından biri
Her iki yöntemde de, ustalar (alet üreticileri), önemli bir bilişsel gelişmeyi temsil edebilecek, istenen ürünü (hiyerarşik organizasyon) yaratmayla dolaylı olarak ilgili bazı öğeler üretmiş olmalıydı. Modern insanlarla yapılan deneyler, platform hazırlığının önceki tekniklerin aksine tamamen gözlemsel öğrenme yoluyla öğrenilemeyeceğini ve kendi kendini düzenleyen öğrenmenin yanı sıra iyi gelişmiş öğretim yöntemlerinin göstergesi olabileceğini göstermiştir. Boxgrove’da knappers, çekiç yapmak için sadece taş değil aynı zamanda kemik ve boynuz da kullandı ve bu kadar geniş bir hammadde yelpazesinin kullanımı, taş işçiliği, çalışmak ve malzeme toplamak için kemik işçiliğinden çok daha farklı bir beceri seti gerektirdiğinden, gelişmiş planlama yetenekleriyle konuşabilirdi.[46]
Kapthurin Formasyonu, Kenya, 545.000 509 yıl önce kadar uzanan bıçak ve dilgicik teknolojinin en eski kanıtlar vermiştir. Bu teknoloji Orta Paleolitik’te bile nadirdir ve tipik olarak Üst Paleolitik modern insanlarla ilişkilendirilir. Bunun uzun bir bıçak yapma geleneğinin parçası olup olmadığı veya bıçak teknolojisinin birden fazla farklı insan türü tarafından birkaç kez kaybolup yeniden icat edilip edilmediği belirsizdir. [48]
ATEŞ KULLANIMI VE İNŞA
Daha soğuk iklimlere yönelik bariz zorlamalara rağmen, arkeolojik kayıtlarda 400 ila 300 bin yıl öncesine kadar ateş kanıtı azdır. Yangın kalıntılarının basitçe bozulmuş olması mümkün olsa da, Arago veya Gran Dolina gibi uzun ve genel olarak bozulmamış işgal sekansları, bariz bir şekilde yangın kullanımına dair ikna edici kanıtlardan yoksundur. Bu model muhtemelen şu anda ateşleme teknolojisinin veya gelişmiş yangın bakım tekniklerinin icadını gösterebilir ve yangın o zamandan önce Avrupa’da insanların yaşamlarının ayrılmaz bir parçası değildi. Öte yandan, Afrika’da insanlar, Afrika’da çok daha sık meydana gelen doğal orman yangınlarından 1,6 milyon yıl kadar önce, sık sık ateşi temizleyebildiler ve bu nedenle muhtemelen (az ya da çok) düzenli olarak ateşi kullandılar. Afrika’nın ötesinde kurulmuş en eski sürekli yangın alanı 780.000 yaşındadır.Gesher Benot Ya’aqov, İsrail. [49]
Avrupa’da, çoğunlukla hava koşullarından korunan alanlarda inşa edilmiş sağlam temellere sahip sağlam yüzeyli kulübeler olarak sınıflandırılan inşa edilmiş konut yapılarının kanıtı, Çek Cumhuriyeti’nin Přezletice kentinden 700.000 yıllık bir taş temelin en eski örneği olan Cromerian Interglacial’dan bu yana kaydedilmiştir. Bu konut muhtemelen kalın dallardan veya ince direklerden yapılmış, büyük kayalar ve topraktan oluşan bir temelle desteklenen tonozlu bir çatıya sahipti. Almanya, Bilzingsleben’de (424.000 yıl önce başlayan) Holstein Interglacial sırasında veya sonrasında var olduğu öne sürülmüştür; Terra Amata, Fransa; ve Fermanville ve Saint-Germain-des-Vaux içinde Normandiya. Bunlar muhtemelen kış aylarında kullanılıyordu ve ortalama olarak sadece 3.5 m × 3 m alandaydılar, muhtemelen sadece uyumak için kullanılıyorlardı, diğer faaliyetler (itfaiye dahil) dışarıda yapılmış gibi görünüyor. Daha az kalıcı çadır teknolojisi, Avrupa’da Aşağı Paleolitik’te mevcut olabilir. [50]
SİLAHLAR
Tekrarlanan ateş kullanımının ortaya çıkışı – Avrupa’da en erken İngiltere’deki Beeches Pit ve Almanya’daki Schöningen’den – kabaca Schöningen mızrakları tarafından en iyi örneklenen saplama teknolojisiyle (taş noktaları mızraklara tutturma) çakışmaktadır.[49] Bu 9 tahta mızrak ve mızrak parçası – bir mızrak ve çift uçlu bir sopaya ek olarak – 300.000 yıl öncesine aittir ve bir göl kenarında korunmuştur. Mızrakların çapları 2,9-4,7 cm arasında değişir ve 210-240 cm (7-8 ft) uzunluğunda olabilir, genel olarak günümüzün rekabetçi ciritlerine benzer. Mızraklar, (aynı zamanda yumuşak) çam ağacı olan mızrak 4 hariç, yumuşak ladin ağacından yapılmıştır. Bu, Clacton öncüsü ile tezat oluşturuyordan Clacton-on-Sea sert yapılmıştı, İngiltere, belki yaklaşık 100.000 yıl daha yaşlı, porsuk ahşap. Schöningen mızrakları 35 m’ye kadar bir menzile sahip olabilir, ancak yaklaşık 5 m içinde daha etkili kısa menzilli olurdu, bu da onları av veya yırtıcılara karşı etkili mesafe silahları haline getirirdi. Bu iki yerleşim yerinin yanı sıra, Avrupa mızrak teknolojisinin sağlam kanıtını sağlayan diğer tek yerleşim yeri, görünüşe göre bir file 238 cm porsuk mızrağının yerleştirildiği 120.000 yıllık Lehringen bölgesi, Almanya’dır.[51] Afrika’da, Güney Afrika’daki Kathu Pan 1’den 500.000 yıllık noktalar mızraklara saplanmış olabilir. Dolaylı kanıtlara göre, bir at kürek kemiği500.000 yıllık Boxgrove’dan bir mızrak yarasıyla uyumlu bir delinme yarası gösteriyor. Hafting kanıtı (hem Avrupa’da hem de Afrika’da) 300.000 yıl sonra çok daha yaygın hale geliyor. [52]
KONUŞMA DİLİ
SH insanları, modern insan benzeri bir dil kemiğine (dili destekleyen) ve normal insan konuşması aralığındaki frekansları ince bir şekilde ayırt edebilen orta kulak kemiklerine sahipti. Diş çizgilerine bakılırsa, ağırlıklı olarak sağlak oldukları görülüyor ve el kullanımı, modern insanlarda tipik olarak dil işleme ile ilişkili olan beyin fonksiyonunun lateralizasyonu ile ilgili. Bu nedenle, bu nüfusun erken bir dil biçimiyle konuştuğu varsayılmaktadır.[45][53][54]Bununla birlikte, bu özellikler, dilin ve insani konuşmanın varlığını kesin olarak kanıtlamaz ve bu tür anatomik tartışmalara rağmen zamanın bu kadar erken zamandaki varlığına öncelikle bilişsel bilim adamı Philip Lieberman karşı çıkmıştır. [53]
KAYNAKÇA
- Harvati, K. (2007). ” Homo heidelbergensis’in 100 yılı – tartışmalı bir taksonun yaşamı ve zamanları” (PDF) Mitteilungen der Gesellschaft für Urgeschichte 16: den 85. Arşivlenmiş orijinal (PDF) 4 Mart 2016 tarihinde.
- Mounier, A.; Martal, F.; Condemi, S. (2009). “Homo heidelbergensisayrı bir tür mü? Mauer çene kemiğine ilişkin yeni anlayış”. İnsan Evrimi Dergisi. 56(3): 219–246. .
- Başladı, David R., ed. (2012). ” Homo sapiens’in Afrika Kökeni “. Paleoantropoloji A Companion. John Wiley ve Oğulları. ISBN’si 9781118332375.
- Schwartz, JH; Tattersall, İ.(2010). “ Homo sapiens’in kökeni için fosil kanıtı“. Amerikan Fiziksel Antropoloji Dergisi. 143(S51): 96–98, 101–103.
- Stringer, CB(2012). “ Homo heidelbergensis’in durumu(Schoetensack 1908)”. Evrimsel Antropoloji. 21(3): 101–104.
- de Lumley, M.-A. (2015). “L’homme de Tautavel. UnHomo erectuseuropéen évolué.Homo erectus tautavelensis“. L’Antropoloji. 119(3): 344-346.
- Buck, LT; Stringer, CB(2014). “ Homo heidelbergensis “. Güncel Biyoloji. 24(6): R214–R215.
- “Hierzu zählte noch im Jahr 2010 auch das Geologisch-Paläontologische Institut Universität Heidelberg der, Unterkiefer den das 1908 verwahrt und ihn als Seit Homo erectus heidelbergensis auswies. İnzwischen Heidelberg als içinde wird er jedoch auchHomo heidelbergensis bezeichnet, Æ” (Almanca) Geowissenschaften için Sammlung des Instituts 29 Kasım 2015’te erişildi
- de Castro, JMB; Martinez, İ.; Gracia-Téllez, A.; Martinon-Torres, M.; Arsuaga, JL (2020). “Sima de los Huesos Orta Pleistosen hominin bölgesi (Burgos, İspanya). Kişi sayısının tahmini”. Anatomik Kayıt. 304 (7): 1463–1477.
- Meyer, M.; Arsuaga, J.-L. de Filippo, C.; et al. (2016). “Orta Pleistosen Sima de los Huesos homininlerinden Nükleer DNA dizileri”. Doğa. 531 (7595): 504–507.
- Stringer, Chris (2012). “Yorum: Modern insanı ne yapar”. Doğa. 485 (7396): 33–35 [34].
- Grün, Rainer; Pike, Alistair; McDermott, Frank; Eggins, Stephen; Mortimer, Graham; Aubert, Maxime; Kinsley, Lesley; Joannes-Boyau, Renaud; Rumsey, Michael; Denis, Christiane; Brink, James; Clark, Tara; Stringer, Chris (1 Nisan 2020). “Broken Hill, Zambiya’dan kafatasının tarihlendirilmesi ve insan evrimindeki konumu” (PDF). Doğa. 580 (7803): 372-375.
- Beyaz, Tim D. Asfaw, B.; DeGusta, D.; Gilbert, H.; Richards, GD; Suwa, G.; Howell, FC (2003). ” Orta Awash, Etiyopya’dan Pleistosen Homo sapiens “. Doğa. 423 (6491): 742-747.
- Asfaw, Berhane (2005). “Bodo, orta Awash Vadisi, Etiyopya’dan yeni bir hominid parietal”. Amerikan Fiziksel Antropoloji Dergisi. 61 (3): 367-371.
- Rightmire, G. Philip (2005). “Göl Ndutu kafatası ve Afrika’da erken Homo sapiens “. Amerikan Fiziksel Antropoloji Dergisi. 61 (2): 245–254.
- Profico, A.; di Vincenzo, F.; et al. (2016). “Boşluğu doldurma. Gombore II’den (Melka Kunture, Etiyopya; yaklaşık 850 bin yıl) insan kafatası kalıntıları ve Homo heidelbergensis’in kökeni“(PDF). Antropolojik Bilimler Dergisi. 94(94): 1-24.
- de Castro, JMB; Martinon-Torres, M.; Arsuaga, JL; Carbonell, E. (2017). ” Homo antecessor’un yirminci yıldönümü (1997-2017): bir inceleme”. Evrimsel Antropoloji. 26 (4): 164.
- Roksandic, Mirjana; Radović, Predrag; Lindal, Joshua (Şubat 2018). ” Homo heidelbergensis’in hipodigmasının gözden geçirilmesi Doğu Akdeniz’den bir görünüm”. Kuvaterner Uluslararası 466: 66-81.
- Rightmire, G. Philip (1998). “Orta Pleistosen’de İnsan Evrimi: Homo heidelbergensis’in Rolü ” (PDF) Evrimsel Antropoloji 6 (6): 218–227.
- Meyer, M.; Arsuaga, J.; de Filippo, C.; Nagel, S. (2016). “Orta Pleistosen Sima de los Huesos homininlerinden Nükleer DNA dizileri”. Doğa 531 (7595): 504–507.
- Bermudez de Castro, JM; Arsuaga, JL; Carbonell, E.; Rosas, A.; Martinez, İ.; Camira, M. (1997). “Atapuerca, İspanya’nın Alt Pleistoseninden Bir Hominid: Neandertallerin ve Modern İnsanların Olası Atası” (PDF) Bilim. 276 (5317): 1392-1395. S2CID 31088294. Arşivlenmiş orijinal (PDF) 2020-02-07 tarihinde.
- de Castro, J.-MB (23 Mayıs 2015). ” Homo antecessor: On sekiz yıl sonraki son teknoloji”. Kuvaterner Uluslararası 433: 22–31.
- Welker, F.; Ramos-Madrigal, J.; Gutenbrunner, P.; et al. (2020-04-01). “Homo antecessor’un diş proteomu” Doğa 580 (7802): 235-238.
- Hosfield, R.; Cole, J. (2018). “Kuzeybatı Avrupa’daki erken homininler: Noktalı uzun bir kronoloji mi?” Kuvaterner Bilim İncelemeleri 190: 148-160.
- Rightmire, RP (2004). “Erken Orta PleistosenHomo’dabeyin büyüklüğü ve ensefalizasyon“. Amerikan Fiziksel Antropoloji Dergisi. 124(2): 109–123.
- Allen, JS; Damasio, H.; Grabowski, TJ (2002). “İnsan beynindeki normal nöroanatomik varyasyon: MRI-hacimsel bir çalışma” Amerikan Fiziksel Antropoloji Dergisi. 118 (4): 341–358.
- Stringer, C. (1984). “Pleistosen’de insan evrimi ve biyolojik adaptasyon”. Gelen Foley, R.(Ed.). Hominid evrimi ve topluluk ekolojisi. Akademik Basın. ISBN’si 978-0-12-261920-5.
- Holloway, RL (1985). ” Homo sapiens neanderthalensis’in zavallı beyni: ne istersen gör”. Delson’da, E. (ed.). Atalar: Sert kanıt. Alan R.Liss. ISBN’si 978-0-471-84376-4.
- Amano, H.; Kikuchi, T.; Morita, Y.; Kondo, Ö.; Suzuki, H.; et al. (2015). “Neandertal Amud 1 Kafatasının Sanal Yeniden İnşası” (PDF). Amerikan Fiziksel Antropoloji Dergisi 158(2): 185–197.
- Carretero, José-Miguel; Rodriguez, Laura; Garcia-González, Rebeca; Arsuaga, Juan-Luis; Gomez-Olivencia, Asier; Lorenzo, Carlos; Bonmati, Alejandro; Gracia, Ana; Martínez, Ignacio (2012). “Sima de los Huesos, Sierra de Atapuerca (İspanya)’dan Orta Pleistosen insanlarındaki tam uzun kemiklerden boy tahmini” (PDF). İnsan Evrimi Dergisi. 62 (2): 242–55.
- Churchill, SE; Berger, LR Hartstone-Rose, A.; Zondo, BH “Afrika, Orta Pleistosen Vücut büyüklüğü Homo“. Afrika Genesis 325-326.
- DeCroos, FC; Liao, JC; Ramey, NA; Li, I (2011-08-15). “Odontojenik Orbital Selülit Yönetimi”. Tıp ve Yaşam Dergisi. 4 (3): 314–317. ISSN 1844-122X. PMC 3168817. PMID 22567060
- Ascaso, F.; Adiego, MI (2016-09-14). “Homo heidelbergensis: en eski odontojenik orbital selülit vakası?”. Acta Oftalmolojik. 94
- Gracia-Téllez, Ana; Arsuaga, Juan-Luis; Martinez, Ignacio; Martín-Frances, Laura; Martinon-Torres, Maria; Bermúdez De Castro, José-María; Bonmati, Alejandro; Lira, Jaime (2013-05-08). “Homo heidelbergensis’te orofasiyal patoloji: Sima de los Huesos sitesinden (Atapuerca, İspanya) Kafatası 5 olgusu”. Kuvaterner Uluslararası. 295: 83-93.
- Bonmati, A.; Gomez-Olivencia, A.; Arsuaga, JL; et al. (2010). “İspanya, Sima de los Huesos bölgesinden yaşlı bir insan bireyinden Orta Pleistosen bel ve pelvis” Ulusal Bilimler Akademisi Bildirileri. 107 (43): 18386-18391
- Garcia, A.; Arsuaga, JL; Martinez, İ.; et al. (2009). “Sima de los Huesos, Atapuerca, İspanya’dan Orta Pleistosen insan Cranium 14’te Kraniosinostoz” Ulusal Bilimler Akademisi Bildirileri 106 (16): 6573-6578.
- Bermudez de Castro, JM; Perez, PJ (1995). “Atapuerca’dan (İspanya) Orta Pleistosen Hominidlerinde Emaye Hipoplazisi”. Amerikan Fiziksel Antropoloji Dergisi. 96 (3): 301–314.
- Isaac, G. (1971). “Erken insanın diyeti: Afrika’daki alt ve orta Pleistosen bölgelerinden arkeolojik kanıtların yönleri”. Dünya Arkeolojisi. 2 (3): 279–299.
- Bigga, G.; Schoch, WH; Kentsel, B. (2015). “Schöningen (Almanya) Orta Pleistosen’de paleoortam ve bitki sömürüsü olanakları. Botanik makro kalıntıları ve polenden içgörüler”. İnsan Evrimi Dergisi. 89: 92-104
- Majkic, A.; d’Errico, F.; Stepanchuk, V. (2018). “Paleolitik oyulmuş kortekslerin öneminin değerlendirilmesi. Kırım, Kiik-Koba’nın Mousterian bölgesinden bir vaka çalışması”. PLOS BİR. 13 (5): e0195049.
- Bednarik, RG (2005). “Acheulian boncukları hakkında daha fazlası” (PDF) Kaya Sanatı Araştırması. 22 (2): 210–212. (Kolaylık bağlantısı)
- Rigaud, S.; d’Errico, F.; Vanhaeren, M.; Neumann, C. (2009). “Varsayılan Acheulean Porosphaera globularis boncuklarının kritik yeniden değerlendirilmesi “. Arkeoloji Bilimi Dergisi. 36 (1): 25–34
- Wreschner, EE; Bolton, R.; et al. (1980). “Kızıl Okra ve İnsan Evrimi: Bir Tartışma Örneği”(PDF) Güncel Antropoloji 21 (5): 631-632.
- Carbonell, E.; Camira, M. (2006). “Sembolik bir davranışın ortaya çıkışı: Sima de los Huesos, Sierra de Atapuerca, Burgos, İspanya’nın mezar çukuru”. Comptes Rendus Palevol. 5(1–2): 155–160.
- Stout, D.; Apel, J.; Komutan, J.; Roberts, M. (2014). “Boxgrove, İngiltere’de Geç Aşölyen teknolojisi ve biliş”. Arkeoloji Bilimi Dergisi. 41: 576-590.
- Dennel, R. (2020). “Movius Hattı olmadan Yaşam: Doğu ve Güneydoğu Asya Erken Paleolitik’in yapısı” Kuvaterner Uluslararası 400: 14–22.
- Johnson, CR; McBrearty, S. (2010). “Kapthurin Formasyonu, Kenya’dan 500.000 yıllık bıçaklar”. İnsan Evrimi Dergisi. 58 (2): 193–200.
- Roebroekes, W.; Villa, P. (2011). “Avrupa’da alışılmış ateş kullanımına ilişkin en eski kanıtlar üzerine”. Ulusal Bilimler Akademisi Bildirileri. 108(13): 5209-5214. Bibcode:2011PNAS..108.5209R.
- Sklenař, K. (1987). “Přezletice’deki Alt Paleolitik Konut Yapısı ve Önemi”. antropoloji. 25(2): 101–103. JSTOR 26294864
- Schoch, WH; Bigga, G.; Böhner, U.; Richter, P.; Terberger, T. (2015). “Schöningen’in Paleolitik bölgesinden ahşap silahlar hakkında yeni görüşler”. İnsan Evrimi Dergisi 89 214-225.
- Wilkins, Jayne; Schoville, Benjamin J.; Brown, Kyle S.; Hazan, Michael (2012). “Erken Hafted Avcılık Teknolojisine İlişkin Kanıtlar” (PDF) Bilim. 338 (6109): 942–6.
- Martínez, I.; Arsuaga, JL; Kum, R.; et al. (2008). “Sima de los Huesos’un (Sierra de Atapuerca, İspanya) orta Pleistosen bölgesinden insan hyoid kemikleri”(PDF). İnsan Evrimi Dergisi. 54(1): 118–124.
- Lozano, Marina; Camii, Marina; De Castro, José María Bermúdez; Arsuaga, Juan Luis; Carbonell, Eudald (2009). ” 500.000 yıl önce Sima de los Huesos’tan (Atapuerca, İspanya) Homo heidelbergensis’in sağlaklığı”. Evrim ve İnsan Davranışı. 30 (5): 369-76.
