Solunum Sistemi

| Solunum sistemi | |
|---|---|
İnsan solunum sistemine parçaları ve işlevleri ile eksiksiz, şematik bir bakış. |
|
| ayrıntılar | |
| Tanımlayıcılar | |
| Latin | solunum sistemi |
| MeSH | D012137 |
| TA | A06.0.00.000 |
| FMA | 7158 |
Solunum sistemi, biyolojik sistemde bulunan tüm canlıların, yaşamını devam ettirebilmesi için gerçekleştirdiği gaz alışverişinin gerçekleştiği organları ve mekanizmanın tamamıdır.
Bunu gerçekleştiren anatomi ve fizyoloji, organizmanın büyüklüğüne, içinde bulunduğu çevreye ve evrimsel tarihine bağlı olarak büyük ölçüde değişir. Kara hayvanlarının solunum yüzeyi balataları olarak içselleştirilmiş olduğu akciğerleridir . [1] Akciğerlerdeki gaz değişimi alveol adı verilen milyonlarca küçük hava kesesinde meydana gelmektedir. Sürüngen ve kuşlarda bu eylem atriyumda gerçekleşir . Bu mikroskobik hava keseleri çok zengin bir kan kaynağına sahiptir, böylece havayı kanla yakın temasa sokar. [2] Bu hava keseleri dış çevre ile bir hava yolları sistemi veya en büyüğü olan göğsün ortasında iki ana bronşın içine giren trakea olan içi boş tüpler yoluyla iletişim kurar . Bunlar akciğerlere girerek gittikçe daha dar olan ikincil ve üçüncül bronşlara girer ve çok sayıda küçük tüpe, bronşiyollere ayrılır . Kuşlarda ise parabronchi denilen bronşiollerde gerçekleşir ve genellikle mikroskobik olarak açılan bronşiyoller veya parabronchi’dir. Hava, solunum kaslarını içeren solunum işlemi ile ortamdan alveollere veya atriyumlara pompalanmalıdır .
Omurgalı ya da omurgasız çoğu balık ve diğer bir dizi su hayvanlarında solunum sistemi solungaçlardan oluşur. Gaz alış verişi bu su solungaçların üzerinde çeşitli aktif veya pasif yollarla akar. Gaz değişimi, ince veya çok düz filamentlerden ve çok vaskülerize edilmiş dokunun çok geniş bir yüzey alanını suya maruz bırakan lammelalardan oluşan solungaçlarda gerçekleşir .
Böcekler gibi diğer hayvanlarda çok basit anatomik özelliklere sahip solunum sistemleri vardır ve amfibilerde cildin gaz alışverişinde bile hayati bir rolü vardır. Bitkiler ayrıca solunum sistemlerine sahiptir, ancak gaz değişiminin yönü hayvanlardakinin tersi olabilir. Bitkilerdeki solunum sistemi, bitkinin çeşitli bölgelerinde bulunan stoma gibi anatomik özellikleri içerir . [3]
Memeliler
Anatomi

Şekil 1. Solunum sistemi
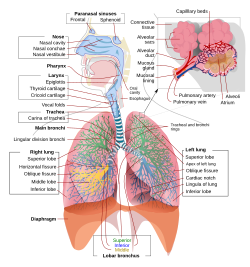
Şek. 2. alt solunum yolları , veya “Solunum ağacı”
- Nefes borusu
- Ana bronş
- Lobar bronş
- Segmental bronş
- bronşiyol
- Alveolar kanal
- Alveol
İnsanlarda ve diğer memelilerde , tipik bir solunum sistemi anatomisidir solunum yolu olarak, sistem bir üst ve bir alt solunum yoluna ayrılmıştır . Üst sistem burun , burun boşlukları , sinüsler , farinks ve gırtlakın vokal kıvrımların üzerindeki kısmını içerir . Alt kanal (Şekil 2.) larinks, trakea , bronşlar , bronşiyoller ve alveollerin alt kısmını içerir .
Alt yolun dallanan hava yolları sıklıkla solunum ağacı veya trakeobronşiyal ağaç olarak tanımlanır (Şekil 2). [4] “Ağaç” ın çeşitli dalları boyunca birbirini takip eden dallanma noktaları arasındaki aralıklara genellikle yetişkin insanda yaklaşık 23 olan dallanma “nesiller” olarak atıfta bulunulmaktadır. Önceki kuşaklar (yaklaşık nesil 0–16), trakea ve bronşların yanı sıra basitçe hava kanalları olarak hareket eden, solunum bronşiyollerine, alveolar kanallara ve alveollere (yaklaşık 17-23 kuşaklar) hava değişimi sağlayan daha büyük bronşiyollerden oluşur (yaklaşık 17-23 kuşaklar) . [5] [6] Bronşiyoller herhangi bir kıkırdak desteği olmayan küçük hava yolları olarak tanımlanmaktadır. [4]
Trakeadan dallanan ilk bronşlar sağ ve sol ana bronşlardır. İkincisi sadece trakea çapı (1.8 cm), bu bronşlar (1 -1.4 cm çapında) [5] her hilumda akciğerlere girer , burada lober bronchi olarak bilinen daha dar ikincil bronşlara dallar ve bu dallar daha dar üçüncül olur. segmental bronşlar olarak bilinen bronşlar. Segmental bronşların (1 ila 6 mm çapında) diğer bölümleri [7] 4. dereceden, 5. dereceden ve 6. dereceden segmental bronşlar olarak bilinir ya da sonradan bronşlar olarak gruplandırılır. [8] [9]
Yetişkin insanda solunum ağacının ortalama 23 dalına kıyasla, farede sadece bu tür 13 dal vardır.
Alveoller, “ağacın” çıkmaz terminalleridir, yani onlara giren havanın aynı yoldan çıkması gerekir. Bunun gibi bir sistem ölü boşluk yaratır , ekshalasyondan sonra hava yollarını dolduran bir hava hacmi (yetişkin insanda yaklaşık 150 ml) ve çevresel hava onlara ulaşmadan önce alveollere geri verilir. [10] [11] hava yolu gaz değiştirici ile temas etmeden solunumla çevre hava ile doldurulur inhalasyon sonunda. [10]
Havalandırma hacimleri
Akciğerler solunum döngüsü sırasında genişler ve büzülür, akciğerlere hava girer ve çıkar. Normal dinlenme koşulları altında akciğerlerin içine veya dışına taşınan hava hacmi (yaklaşık 500 ml’lik dinlenme gelgit hacmi ) ve maksimum zorla inhalasyon ve maksimum zorla ekshalasyon sırasında taşınan hacimler, spirometri ile ölçülür . [12] Tipik bir yetişkin insan spirogramı, akciğerlerin çeşitli hacimlerde gezilere verilebildiği isimlerle aşağıda gösterilmiştir (Şekil 3):

Şekil 3 Bir ‘spirometrenin’ çıkışı. Grafiğin soldan okunan yukarı doğru hareketi hava girişini gösterir; aşağı hareketler ekshalasyonu temsil eder.
Maksimum zorla ekshalasyon sırasında akciğerlerdeki tüm hava dışarı atılmaz. Bu, spirometri ile ölçülemeyen yaklaşık 1.0-1.5 litrelik kalıntı hacmidir . Kalıntı hacmi (yani yaklaşık 2.5-3.0 litre fonksiyonel kalıntı kapasitesi ve yaklaşık 6 litre toplam akciğer kapasitesi) içeren hacimler de spirometri ile ölçülemez. Ölçümleri özel teknikler gerektirir. [12]
Havanın ağızdan veya burundan veya alveollerin içine veya dışına soluma hızları, nasıl hesaplandıklarıyla birlikte aşağıdaki tabloda verilmiştir. Dakikadaki nefes döngüsü sayısı solunum hızı olarak bilinir .
| Ölçüm | Denklem | Açıklama |
|---|---|---|
| Dakika havalandırma | tidal hacim * solunum hızı | dakikada burun veya ağza giren veya çıkan toplam hava hacmi. |
| Alveolar ventilasyon | (gelgit hacmi – ölü boşluk) * solunum hızı | dakikada alveollere giren veya çıkan havanın hacmi. |
| Ölü alan havalandırması | ölü boşluk * solunum hızı | soluma sırasında alveollere ulaşmayan, ancak dakika başına hava yollarında kalan hava hacmi. |
Solunum mekaniği

6 Solunum sırasında göğüs kafesinin göğüs hareketlerinin gerçek zamanlı manyetik rezonans görüntüleme (MRI)

Şekil 4 Göğüs kafesinin genişlemesinde solunum kaslarının etkisi . Burada gösterilen özel eylem , göğüs kafesinin pompa kolu hareketi olarak adlandırılır .

Şekil 5 Göğüs kafesinin bu görünümünde, orta hattan dışa doğru alt nervürlerin aşağı doğru eğimi açıkça görülebilir. Bu, “pompa kolu etkisine” benzer bir harekete izin verir, ancak bu durumda kova kolu hareketi olarak adlandırılır .
Nefes

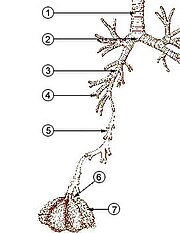
Şekil 8 Kuvvetli solunum kasları (inhalasyon ve ekshalasyon). Renk kodu soldaki ile aynıdır. Diyaframın daha kuvvetli ve kapsamlı bir şekilde kasılmasına ek olarak, interkostal kaslara, kaburgaların hareketini yukarı doğru abartmak için solunum inhalasyon kasları yardımcı olur ve göğüs kafesinin daha büyük bir genişlemesine neden olur. Ekshalasyon sırasında, inhalasyon kaslarının gevşemesinin yanı sıra, karın kasları göğüs kafesinin alt kenarlarını aşağıya doğru çekmek için aktif olarak kasılırken, göğüs kafesinin hacmini azaltırken, aynı zamanda diyaframı toraksın derinliklerine doğru iter.
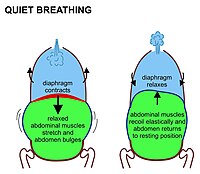
Olarak , memelilerde , istirahat inhalasyon kasılması başta gelen nedeni, diyafram. Bu, torasik boşluğu karın boşluğundan ayıran yukarı doğru kubbeli bir kas tabakasıdır. Tabaka kasıldığında torasik boşluğun hacmini artırarak düzleşir (yani Şekil 7’de gösterildiği gibi aşağı doğru hareket eder). Kasılma diyaframı karın organlarını aşağı doğru iter. Ancak pelvik taban, en alt karın organlarının bu yönde hareket etmesini önlediğinden, esnek karın içerikleri, göbeğin öne ve yanlara doğru dışarı çıkmasına neden olur, çünkü rahat karın kasları bu harekete direnmez (Şekil 7). Normal solunum sırasında karnın bu tamamen pasif şişkinliği (ve ekshalasyon sırasında büzülmesi), bazen vücudun dışında görünmeyen “diyafragmatik solunum” olarak adlandırılır. Memeliler karın kaslarını sadece güçlü ekshalasyon sırasında kullanırlar (bkz.Şekil 8 ve aşağıdaki tartışma). Asla herhangi bir inhalasyon sırasında.
Diyafram kasıldığında, göğüs kafesi Şekil 4’te gösterildiği gibi kaburgaların interkostal kaslar tarafından yukarı doğru çekilmesi ile aynı anda büyütülür ( Tüm kaburgalar arkadan öne doğru eğiktir (Şekil 4’te gösterildiği gibi); ancak en alttaki kaburgalar da orta hattan dışa doğru aşağı doğru eğimlidir (Şek. 5). Böylece, göğüs kafesinin enine çapı, Şekil 4’te gösterilen pompa kolu hareketi ile antero-posterior çap arttıkça arttırılabilir .
Torasik boşluğun dikey boyutunun diyaframın kasılmasıyla genişlemesi ve iki yatay boyutu, kaburgaların ön ve yanlarının kaldırılmasıyla, toraks içi basıncın düşmesine neden olur. Akciğerlerin iç kısımları dış havaya açıktır ve elastiktir, bu nedenle artan alanı doldurmak için genişler. Akciğerlere hava girişi solunum solunum yolları yoluyla gerçekleşir (Şekil 2). Sağlıkta bu hava yolları (burun veya ağızdan başlayıp alveol adı verilen mikroskopik çıkmaz keselerde biten ) her zaman açıktır, ancak çeşitli bölümlerin çapları sempatik ve parasempatik sinir sistemleri tarafından değiştirilebilir.. Bu nedenle alveolar hava basıncı, istirahat halindeyken atmosferik hava basıncına ( deniz seviyesinde yaklaşık 100 kPa ) yakındır ve solunum sırasında havanın akciğerlere girip çıkmasına neden olan basınç gradyanları nadiren 2-3 kPa’yı aşar. [13] [14]
Ekshalasyon sırasında diyafram ve interkostal kaslar gevşer. Bu, göğsü ve karnı anatomik elastikiyetleri ile belirlenen bir konuma döndürür. Akciğerler , yetişkin insanda fonksiyonel kalıntı hava kapasitelerini (Şekil 7’nin sağ elindeki açık mavi alan ) içerdiğinde, göğüs ve karnın “dinlenme orta konumu” dur (Şekil 7) . yaklaşık 2.5-3.0 litre hacme sahiptir (Şekil 3). [6] Dinlenme ekshalasyonu inhalasyondan yaklaşık iki kat daha uzun sürer, çünkü diyafram inhalasyon sırasında aktif olarak kasıldığından daha pasif bir şekilde gevşer.

Şekil 9 İstirahat halindeyken normal bir solunum döngüsü sırasında alveolar havanın bileşimindeki değişiklikler. Soldaki ve mavi çizgi üzerindeki ölçek, kPa’daki kısmi karbondioksit basınçlarını gösterirken, sağdaki ve kırmızı çizgi, kPa’daki de oksijen kısmi basınçlarını gösterir (kPa’yı mm Hg’ye dönüştürmek için, çarpın 7.5).
Tek bir solunum döngüsü sırasında (burun veya ağızda) içeri veya dışarı hareket eden hava hacmine gelgit hacmi denir . Dinlendirici bir yetişkin insanda, nefes başına yaklaşık 500 ml’dir. Ekshalasyonun sonunda hava yolları, inhalasyon sırasında alveollere geri solunan ilk hava olan yaklaşık 150 ml alveolar hava içerir. [10] [15] alveol nefesle ve arka yeniden olarak da bilinen bu hacimli hava ölü boşluk doğurur havalandırma, bu 500 mi her nefeste alveollere sadece 350 ml (500 ml nefes arasında – 150 ml = 350 ml) taze ılık ve nemlendirilmiş havadır. [6]Bu 350 ml temiz hava, normal ekshalasyondan sonra alveollerde kalan hava ile iyice karıştırıldığı ve seyreltildiği için (yani yaklaşık 2.5-3.0 litre fonksiyonel kalıntı kapasitesi ), alveolar havanın bileşiminin çok az değiştiği açıktır. solunum döngüsü sırasında (bkz.Şekil 9). Oksijen gerilimi (veya kısmi basınç) 13-14 kPa’ya (yaklaşık 100 mm Hg) ve karbon dioksitin 5.3 kPa’ya (veya 40 mm Hg) çok yakın kalır. Bu, denizdeki kuru dış havanın, kısmi oksijen basıncının 21 kPa (veya 160 mm Hg) ve karbon dioksit 0.04 kPa (veya 0.3 mmHg) olduğu bileşimle zıttır. [6]
Ağır solunum sırasında ( hiperpne ), örneğin egzersiz sırasında, inhalasyon, büzülme diyaframının dinlenmeye göre daha güçlü ve daha büyük bir gezintisi ile gerçekleşir (Şekil 8). Ek olarak ” inhalasyon yardımcı kasları “, interkostal kasların hareketlerini abartır (Şek. 8). Bu aksesuar inhalasyon kasları, servikal omurlardan ve kafatasının tabanından üst kaburgalara ve sternuma , bazen klavikulalara bir ara bağlanma yoluyla uzanan kaslardır . [6]Göğüs kafesi kasıldığında, sadece interkostal kasların kasılmasıyla elde edilebilecek seviyeden çok daha fazla artar. Vücudun dışından bakıldığında, yorucu veya zor çalışan inhalasyon sırasında klavikulaların kaldırılmasına bazen özellikle astım atakları sırasında ve kronik obstrüktif akciğer hastalığı olan kişilerde görülen klaviküler solunum denir .
Ağır nefes alma sırasında, ekshalasyon, tüm inhalasyon kaslarının gevşemesinden kaynaklanır. Ancak şimdi, karın kasları, rahat kalmak yerine (istirahat ettikleri gibi), göğüs kafesinin alt kenarlarını aşağı doğru (ön ve yanlar) aşağı doğru çekerek büzülür (Şek. 8). Bu sadece göğüs kafesinin büyüklüğünü büyük ölçüde azaltmakla kalmaz, aynı zamanda karın organlarını yukarı doğru toraks içine doğru şişen diyaframa doğru iter (Şekil 8). Ekshalasyon sonu akciğer hacmi artık dinlenme orta konumunun çok altındadır ve dinlenme “fonksiyonel rezidüel kapasitesinden” çok daha az hava içerir. Bununla birlikte, normal bir memelide akciğerler tamamen boşaltılamaz. Yetişkin bir insanda, maksimum ekshalasyondan sonra akciğerlerde her zaman hala en az 1 litre artık hava kalır. [6]
Otomatik ritmik nefes alıp verme, öksürme, hapşırma (çok güçlü ekshalasyon formları), çok çeşitli duyguların ifadesi (gülme, iç çekme, acı içinde ağlama, bıkkın nefes alımları) ve bu şekilde kesilebilir. konuşma, şarkı söyleme, ıslık çalma ve üflemeli çalgıların çalınması gibi gönüllü eylemler. Tüm bu eylemler yukarıda açıklanan kaslara ve akciğerlerin içindeki ve dışındaki havanın hareketi üzerindeki etkilerine dayanır.
Bir solunum şekli olmamasına rağmen, Valsalva manevrası solunum kaslarını içerir. Bu, aslında, sıkıca kapatılmış karşı çok güçlü exhalatory çaba glottisin hava akciğerlerden kaçabilir böylece. [16] Bunun yerine karın içeriği pelvik tabandaki deliklerden ters yönde boşaltılır. Karın kasları çok güçlü bir şekilde kasılır, karın ve toraks içindeki basıncın aşırı yüksek seviyelere çıkmasına neden olur. Valsalva manevrası gönüllü olarak yapılabilir, ancak daha genel olarak, örneğin zor dışkılama veya doğum sırasında karnı boşaltmaya çalışırken ortaya çıkan bir reflekstir. Bu manevra sırasında solunum durur.
Gaz takası
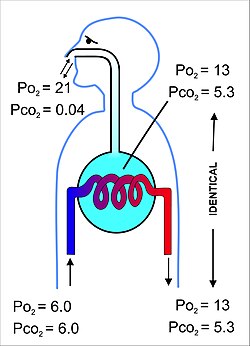
Şekil 11 Memeli akciğerlerindeki gaz değişim sürecinin, ortam havasının gaz bileşimleri, pulmoner kılcal kanın dengelendiği alveolar hava (açık mavi) ve kan gazı gerilimleri arasındaki farkları vurgulayan oldukça şematik bir çizim pulmoner arterde (soldaki akciğere giren mavi kan) ve venöz kanda (kırmızı kan akciğeri sağda bırakır). Tüm gaz gerilimleri kPa cinsindendir. Mm Hg’ye dönüştürmek için 7,5 ile çarpın.

Şekil 12 Akciğer dokusunun normal olarak şişirilmiş bir alveolus (normal ekshalasyonun sonunda) gösteren bir kısmı ve pulmoner kılcal damarları içeren duvarları (kesitte gösterilmiştir ) gösteren diyagramatik bir histolojik kesit . Bu, pulmoner kılcal kanın tamamen alveolar hava ile nasıl çevrildiğini gösterir. Normal bir insan akciğerinde tüm alveoller birlikte yaklaşık 3 litre alveolar hava içerir. Tüm pulmoner kılcal damarlar yaklaşık 100 ml kan içerir

Şekil 10 Gazların kan plazması ve alveolar hava arasında hareket etmesi gereken katmanları gösteren bir alveolar duvardan alınan histolojik bir kesit. Koyu mavi nesneler, kılcal endotelyal ve alveolar tip I epitel hücrelerinin (veya tip 1 pnömositlerin ) çekirdeğidir . “RBC” etiketli iki kırmızı nesne , pulmoner kapiller kandaki kırmızı kan hücreleridir .
Solunum sisteminin birincil amacı, alveolar havadaki solunum gazlarının kısmi basınçlarının pulmoner kılcal kandaki basınçlarla dengelenmesidir (Şekil 11). Bu işlem, basit bir cereyan difüzyon , [17] (olarak bilinen çok ince bir membran boyunca kan hava bariyeri duvarlarını oluşturan), akciğer alveollerinin (Şek. 10). Bu oluşan alveolar epitel hücrelerinin , bunların temel zarlarda ve endotel hücreleri alveoler kılcal (Şek. 10). [18] Bu kan gazı bariyeri son derece incedir (insanlarda ortalama 2,2 μm kalınlığında). Yaklaşık 300 milyon küçük hava kesesine katlanır.alveoller [18] , solunum ile ilgili kollara (çapı ise 75 ila 300 um) bronşiyollerde olarak akciğer , böylece çok büyük bir yüzey alanına (yaklaşık 145 m sağlayan 2 gaz değişimi meydana gelmesi için). [18]
Alveollerin içinde bulunan hava, alveolar kılcal kanı tamamen çevreleyen yaklaşık 2.5-3.0 litrelik yarı kalıcı bir hacme sahiptir (Şekil 12). Bu, iki bölmedeki gazların kısmi basınçlarının dengelenmesinin çok verimli ve çok hızlı gerçekleşmesini sağlar. Alveolar kılcal damarları terk eden ve sonunda vücuda dağıtılan kanın kısmi oksijen basıncı 13-14 kPa (100 mmHg) ve kısmi karbon dioksit basıncı 5,3 kPa (40 mmHg) (yani alveollerde olduğu gibi oksijen ve karbon dioksit gazı gerilimleri). [6] Yukarıdaki bölümde belirtildiği gibideniz ortamındaki (kuru) havadaki oksijen ve karbondioksitin karşılık gelen kısmi basınçları sırasıyla 21 kPa (160 mmHg) ve 0.04 kPa’dır (0.3 mmHg). [6]
Alveolar havanın bileşimi ile ortam havasının bileşimi arasındaki bu belirgin fark korunabilir, çünkü fonksiyonel artık kapasite oldukça dar ve nispeten uzun tüpler (hava yolları: burun , yutak) ile dış havaya bağlanan çıkmaz keselerde bulunur. , larinks , trakea , bronşlar ve dalları bronşiyollere kadar), içinden havanın hem içeri hem de dışarı solunması gerekir (yani kuş akciğerinde olduğu gibi tek yönlü bir akış yoktur)). Bu tipik memeli anatomisi, akciğerlerin her nefesle boşaltılmaması ve yeniden şişirilmemesi (ekshalasyondan sonra alveollerde yaklaşık 2.5-3.0 litre önemli miktarda hava bırakarak) alveolların kompozisyonunun Her inhalasyon ile içine 350 ml temiz hava karıştırıldığında hava sadece minimum düzeyde bozulur. Böylece hayvana, bileşimi günümüzdeki ortam havasından önemli ölçüde farklı olan çok özel bir “taşınabilir atmosfer” sağlanır . [19] Kanın ve dolayısıyla vücut dokularının maruz kaldığı bu taşınabilir atmosfer ( fonksiyonel rezidüel kapasite ) – dış havaya değil.
Ortaya çıkan arteriyel kısmi oksijen ve karbondioksit basınçları homeostatik olarak kontrol edilir . CO arteriyel parsiyel basıncında bir artış 2 ve daha az bir ölçüde, O arteriyel parsiyel basıncında bir düşüş 2 , reflexly daha derin ve daha hızlı kadar nefes neden olur , kan gazı ölçümleri ve akciğerlerde arteriel kan normale dön. Bunun tersine, karbondioksit gerginliği düştüğünde ya da daha az bir ölçüde oksijen gerginliği arttığında gerçekleşir: kan gazı normallerine dönülene kadar solunum hızı ve derinliği azalır.
Alveolar kılcal gelen kan O kısmi basınca sahip yana 2 ortalama bölgesinin, 6 kPa (45 mm Hg), alveoler hava basınç 13-14 kPa (100 mmHg), bir ağ difüzyon olacak iken 3 litre alveolar havanın bileşimini hafifçe değiştirerek kılcal kanın içine oksijen. Benzer şekilde, alveoler kılcal damarlara gelen kanın kısmi bir CO 2 basıncı olduğu içinalveolar havanın 5.3 kPa (40 mmHg) olduğu halde, kılcal damarlardan alveollere doğru net bir karbondioksit hareketi vardır. Bu bağımsız münferit gazların alveolar havaya girip çıkmasıyla meydana gelen değişiklikler, alveolar havanın yaklaşık% 15’inin her 5 saniyede bir ortam havası ile değiştirilmesini gerektirir. Bu çok sıkı bir şekilde göre (doğru alveol hava bileşimini yansıtmaktadır) arteryel kan gazları izlenmesi ile kontrol edilir aort ve karotid organları yanı sıra ile kan gazı ve pH sensörü ön yüzeyinde medula oblongatabeyinde. Akciğerlerde oksijen ve karbondioksit sensörleri de vardır, ancak bunlar öncelikle bronşiyollerin ve pulmoner kılcal damarların çaplarını belirler ve bu nedenle hava ve kan akışını akciğerlerin farklı bölgelerine yönlendirmekten sorumludur.
Sadece 3 litre alveolar havanın bileşiminin doğru bir şekilde korunmasının bir sonucu olarak, her nefesle atmosfere bir miktar karbon dioksit deşarj edilir ve dış havadan bir miktar oksijen alınır. Kısa bir hiperventilasyon periyodu ile normalden daha fazla karbondioksit kaybedilirse , alveolar kısmi karbondioksit basıncı 5.3 kPa’ya (40 mmHg) dönene kadar solunum yavaşlatılacak veya durdurulacaktır. Bu nedenle, solunum sisteminin birincil işlevinin vücudu karbondioksit “atıklarından” kurtarmak olduğu kesin değildir. Her nefesle solunan karbon dioksit muhtemelen vücudun hücre dışı sıvı karbon dioksit ve pH homeostatlarının bir yan ürünü olarak görülebilir.
Bu homeostatlardan ödün verilirse, bir solunum asidozu veya bir solunum alkalozu meydana gelecektir. Uzun vadede bu renal ayarlamalar ile telafi edilebilir , H + ve HCO 3 – plazmadaki konsantrasyonları ; ancak bu zaman aldığından, hiperventilasyon sendromu , örneğin, ajitasyon veya anksiyete, bir kişinin hızlı ve derin nefes almasına neden olduğunda, böylece kandan dış havaya çok fazla CO 2 üfleyerek rahatsız edici bir solunumsal alkaloza neden olduğunda ortaya çıkabilir . [20]
Oksijen suda çok düşük bir çözünürlüğe sahiptir ve bu nedenle hemoglobin ile birlikte gevşek bir şekilde kanda taşınır . Oksijen, hemoglobin molekülü başına dört demir içeren demir içeren hem grubu tarafından hemoglobin üzerinde tutulur . Tüm hem grupları bir O taşıdığınızda 2 molekülü, her kan oksijen ile “doymuş” olduğu söylenir ve oksijen kısmi basıncında ilave bir artış anlamlı kanın oksijen konsantrasyonu artacaktır. Kandaki karbon dioksitin çoğu plazmada bikarbonat iyonları (HCO 3 – ) olarak taşınır . Ancak çözülmüş CO dönüşümü 2 HCO içine 3 –(su ilavesiyle), kanın bir yandan dokulardan, diğer yandan alveolar kılcal damarlardan dolaşım hızı için çok yavaştır. Bu nedenle reaksiyon, kırmızı kan hücrelerinin içindeki bir enzim olan karbonik anhidraz ile katalize edilir . [21] , reaksiyon CO hakim olan kısmi basıncına bağlı olarak her iki yönde olabilir 2 . [6] Hemoglobin moleküllerinin protein kısmı üzerinde karbamino grupları olarak az miktarda karbondioksit taşınır . CO çözülmüş bikarbonat iyonlarının biçimde karbon dioksit toplam konsantrasyonu ( 2ve karbamino grupları) (örn. alveolar havayla dengelendikten sonra) doymuş arteriyel kandaki oksijen konsantrasyonu ile karşılaştırıldığında yaklaşık 22 mM (veya 58 ml / 100 mi) [22] ‘dir. veya 20 ml / 100 ml kan). [6]
Havalandırma kontrolü
Memelilerde akciğer havalandırma yoluyla gerçekleşir nefes alma merkezleri olarak medula oblongata ve pons ve beyin sapı . [6] Bu alanlar, bir dizi oluşturan sinir yolları hakkında bilgi almak , oksijen ve karbon dioksit kısmi basınçlarının içinde arteriyel kan . Bu bilgi , bu basınçları sabit tutmak için akciğerlerin alveollerinin ortalama ventilasyon hızını belirler . Solunum merkezi bunu diyaframı ve diğerlerini harekete geçiren motor sinirler aracılığıyla yapar.solunum kasları .
Kandaki kısmi karbondioksit basıncı arttığında solunum hızı artar. Bu, medulla oblongata’nın ön yüzeyindeki merkezi kan gazı kemoreseptörleri tarafından tespit edilir . [6] aort ve karotid organları , olan periferal kan gazı kemoreseptörler arter özellikle duyarlıdırlar O kısmi basıncı 2 olsa da cevap, ancak daha az güçlü bir şekilde, kısmi basıncına CO 2 . [6]Deniz seviyesinde, normal şartlar altında, solunum hızı ve derinliği, solunum merkezlerinden önce oldukça geniş bir aralık içinde değişmesine izin verilen arteriyel kısmi oksijen basıncı yerine birincil olarak arteriyel kısmi karbon dioksit basıncı ile belirlenir . medulla oblongata ve pons, solunum hızını ve derinliğini değiştirmek için buna cevap verir. [6]
Egzersiz , egzersiz kaslarının gelişmiş metabolizması tarafından üretilen ekstra karbondioksit nedeniyle solunum hızını artırır. [23] uzuvların, katılma pasif hareketleri de refleks solunum oranındaki bir artış meydana getirir. [6] [23]
Akciğerlerdeki streç reseptörlerinden alınan bilgiler gelgit hacmini (inhalasyon ve ekshalasyon derinliği) sınırlar.
Düşük atmosferik basınçlara yanıtlar
Alveoller alveoler hava basıncı yüksekliği için, tam olarak deniz seviyesinde çevre hava basıncı ile aynıdır, ya da herhangi bir suni bir ortamda (örneğin, bir dalış odası veya basınç odası, bu vesile ile atmosfere (solunum yoluyla) açıktır ) bireyin özgürce nefes alması. Akciğerlerin genişlemesi ile (diyaframın indirilmesi ve torasik kafesin genişlemesi yoluyla ) alveolar hava artık daha büyük bir hacme sahiptir ve basıncı orantılı olarak düşerek havanın çevreden, hava yollarından basınca kadar akmasına neden olur. alveoller bir kez daha ortam hava basıncındadır. Bunun tersi, ekshalasyon sırasında açıkça görülür. Bu süreç(teneffüs etme ve ekshalasyon), deniz seviyesinde, Mt. Everest veya bir dalış odası veya dekompresyon odası içinde .
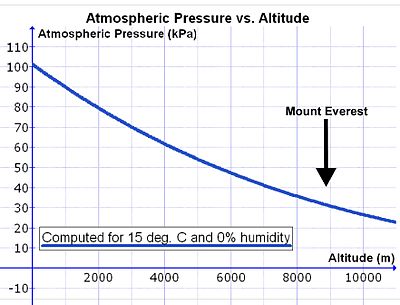
Şekil 14 Toplam atmosfer basıncı ile deniz seviyesinden yükseklik arasındaki ilişkiyi gösteren bir grafik.
Bununla birlikte, deniz seviyesinin üzerine çıktıkça havanın yoğunluğu katlanarak azalır (bkz. Şekil 14) ve yaklaşık 5500 m yükseklikteki her yükselişte yarıya iner . [24] Atmosferik havanın bileşimi, havanın sürekli karıştırma etkisinin bir sonucu olarak neredeyse 80 km’nin altında olduğu için, havadaki oksijen konsantrasyonu ( litre ortam havası başına mmol O 2 ) aynı anda azalır. yükseklikte hava basıncındaki düşüş olarak oran. [25] Bu nedenle, dakikada aynı miktarda oksijen solumak için, kişi deniz seviyesinden daha yüksekte dakika başına orantılı olarak daha fazla hava solumalıdır. Bu daha derin ve daha hızlı nefes alarak elde edilir (yani hiperpne) deniz seviyesinden (aşağıya bakınız).

Şekil 13 Everest Dağı’nın güneyden, Nuptse ve Lhotse’nin arkasında havadan çekilmiş fotoğrafı .
Bununla birlikte, akciğerlere deniz seviyesinde olduğu gibi aynı miktarda oksijen sağlamak için dakika başına solunması gereken hava hacmini ( solunum dakika hacmi ) artıran bir komplikasyon vardır . İnhalasyon sırasında hava, burun pasajlarından ve farenksten geçişi sırasında su buharı ile ısıtılır ve doyurulur . Doymuş su buharı basıncı sadece sıcaklığa bağlıdır. 37 ° C vücut çekirdek sıcaklığında, yükseklik de dahil olmak üzere diğer etkilerden bağımsız olarak 6.3 kPa’dır (47.0 mmHg). [26] Böylece, atmosferik basıncın yaklaşık 100 kPa olduğu deniz seviyesinde, trakeadan akciğerlere akan nemlendirilmiş havasu buharı (6.3 kPa), azot (74.0 kPa), oksijen (19.7 kPa) ve eser miktarda karbon dioksit ve diğer gazlardan (toplam 100 kPa) oluşur. Kuru hava içinde kısmi basıncı -O 2 deniz seviyesinde oksijen 19.7 kPa alveoler hava girmeden kıyasla, 21.0 kPa (100 kPa, yani% 21) ‘dir. (Trakeal kısmi oksijen basıncı [100 kPa – 6,3 kPa] = 19,7 kPa’nın% 21’i kadardır). Mt zirvesinde. Everest (8,848 m veya 29,029 ft yükseklikte) toplam atmosfer basıncı 33,7 kPa’dır ve bunun 7,1 kPa (veya% 21) oksijendir. [24]Akciğerlere giren havanın toplam basıncı 33.7 kPa’dır, bunun 6.3 kPa’sı kaçınılmaz olarak su buharıdır (deniz seviyesinde olduğu gibi). Bu, alveollere giren kısmi oksijen basıncını 5,8 kPa’ya (veya [% 33,7 kPa – 6,3 kPa] = 5,8 kPa’ya) azaltır. Bu nedenle, solunan havadaki kısmi oksijen basıncındaki azalma, toplam atmosfer basıncının irtifadaki azalmasından önemli ölçüde daha fazladır (Everest Dağı’nda: 5.8 kPa’ya karşı 7.1 kPa).
Yükseklikte başka bir küçük komplikasyon daha vardır. Solunum başlangıcında akciğerlerin hacmi anında iki katına çıkarılırsa, akciğerlerin içindeki hava basıncı yarıya iner. Bu yükseklikten bağımsız olarak gerçekleşir. Böylece, deniz seviyesindeki hava basıncının (100 kPa) yarıya indirilmesi, 50 kPa’lık bir intrapulmoner hava basıncına yol açar. Aynı şeyi atmosfer basıncının sadece 50 kPa olduğu 5500 m’de yapmak, intrapulmoner hava basıncı 25 kPa’ya düşer. Bu nedenle, deniz seviyesindeki akciğer hacmindeki aynı değişiklik, ortam havası ile intrapulmoner hava arasındaki basınçta 50 kPa’lık bir fark ile sonuçlanırken, 5500 m’de sadece 25 kPa’lık bir fark ile sonuçlanır. İnhalasyon sırasında havayı akciğerlere zorlayan itici basınç bu nedenle bu yükseklikte yarıya iner. orandeniz seviyesinde soluma sırasında akciğerlere hava girmesi 5500 m. Bununla birlikte, gerçekte, inhalasyon ve ekshalasyon, verilen örneğe göre çok daha nazik ve daha az aniden ortaya çıkar. Solunum döngüsü sırasında akciğerlere hava girip çıkan atmosferik ve intrapulmoner basınçlar arasındaki farklar sadece 2-3 kPa civarındadır. [13] [14] Bu küçük basınç farklılıklarının iki katına veya daha fazlasına ancak yüksek irtifalarda solunum çabasındaki çok büyük değişiklikler sağlanabilir.
Düşük atmosferik basınçların solunum üzerindeki yukarıdaki etkilerinin tümü öncelikle daha derin ve daha hızlı nefes alarak (hiperpne ) karşılanır . Hiperpnenin kesin derecesi , arteriyel kandaki kısmi oksijen ve karbondioksit basınçlarını düzenleyen kan gazı homeostası tarafından belirlenir . Bu homeosta , karbondioksitin arteriyel kısmi basıncının deniz seviyesinde oksijene göre düzenlenmesine öncelik verir . [6] Bu, CO arter kısmi basıncı deniz seviyesinde demek olan 2 O arter kısmi basıncının pahasına, çok yakın 5.3 kPa (ya da 40 mm Hg) üzere geniş bir koşullar aralığı altında tutulur 2düzeltici bir ventilasyon yanıtı ortaya çıkarmadan önce çok geniş bir değer aralığında değişmesine izin verilir. Atmosferik basınç (ve bu nedenle O kısmi basıncı, ancak 2 , havadaki) deniz seviyesinde değeri 50-75 altında% düşer, oksijen homeostazı karbondioksit homeostazı öncelik verilir. [6] Bu geçiş yaklaşık 2500 m (veya yaklaşık 8000 ft) yükseklikte gerçekleşir. Bu anahtar nispeten aniden ortaya çıkarsa, yüksek irtifadaki hiperpne , arteriyel plazmanın pH’ında bir artışla birlikte arteriyel kısmi karbondioksit basıncında ciddi bir düşüşe neden olur . Bu yüksek irtifa hastalığına katkıda bulunanlardan biridir. Öte yandan, oksijen homeostaza geçiş eksikse, hipoksi klinik tabloyu potansiyel olarak ölümcül sonuçlarla karmaşıklaştırabilir.
Daha küçük bronşlarda ve bronşiyollerde oksijen sensörleri vardır . Solunan havadaki düşük kısmi oksijen basınçlarına yanıt olarak, bu sensörler refleksif olarak pulmoner arteriyollerin daralmasına neden olur. [27] (Bu, O 2 düşük arteriyel kısmi basınçlarının arteriyolar vazodilatasyona neden olduğu dokulardaki karşılık gelen refleksin tam tersidir .) Yükseklikte, pulmoner arteriyel basıncın artmasına neden olur ve bu da kan akışının çok daha eşit dağılımına neden olur. deniz seviyesinden daha fazla akciğerlere. Deniz seviyesinde pulmoner arter basıncı çok düşüktür, sonuçta akciğerlerin üstleri bazlardan çok daha az kan alırkan ile nispeten fazla perfüze olan. Alveollerin kan ve hava akışının ideal olarak eşleşmesi sadece akciğerlerin ortasındadır . Yükseklikte , alveollerin akciğerlerin tepelerinden diplere havalandırma / perfüzyon oranındaki bu değişiklik ortadan kaldırılır, tüm alveoller fizyolojik olarak ideal şekilde az çok havalandırılır ve havalandırılır. Bu, yüksek rakımlara ve düşük oksijen basınçlarına uyumlaştırmaya bir diğer önemli katkıdır .
Böbrekler, oksijen ölçmek içeriği (mmol O 2 / litre kan yerine O kısmi basıncı 2 ) arteryel kan. Kanın oksijen muhtevası, yüksek bir irtifada olarak, kronik olarak düşük olduğu zaman, oksijen duyarlı böbrek hücreleri salgılar eritropoietin (genellikle sadece olarak kısaltılmış biçimde bilinen EPO ) [28] kana. [30] Bu hormon, kırmızı kemik iliğini , kırmızı kan hücresi üretim hızını arttırmak için uyarır , bu da kanın hematokritinde bir artışa ve bunun sonucunda oksijen taşıma kapasitesinde (şu anda yüksek hemoglobin nedeniyle) bir artışa neden olur.kan içeriği). O, aynı arter kısmi basıncında, diğer bir deyişle, 2 , yüksek hematokrit olan bir kişinin daha düşük bir hematokrit yapar sahip bir kişiden bir kan litre başına daha fazla oksijen taşır. Yüksek irtifa sakinleri bu nedenle deniz seviyesindeki sakinlerden daha yüksek hematokritlere sahiptir. [29] [30]
Akciğerlerin diğer fonksiyonları
Yerel savunmalar
Nazal pasajlarda veya solunum yollarındaki sinir uçlarının tahrişi, öksürük refleksine ve hapşırmaya neden olabilir . Bu tepkiler havanın sırasıyla trakea veya burundan dışarı atılmasına neden olur . Bu şekilde, solunum yolunu çizen mukusta yakalanan tahriş edici maddeler, yutulabilecekleri yere atılır veya ağız içine taşınır . [6]Öksürme sırasında, hava yolu duvarlarındaki düz kasın kasılması, kıkırdak plakalarının uçlarını birlikte çekerek ve yumuşak dokuyu lümene iterek trakeayı daraltır. Bu, tahriş edici partikül veya mukusu çıkarmak ve çıkarmak için süresi dolmuş hava akış hızını artırır.
Solunum epitelyumu , akciğerlerin savunmasına yardımcı olan çeşitli moleküller salgılayabilir. Bunlar salgı immünoglobulinleri (IgA), koleksiyonlar , defensinler ve diğer peptitler ve proteazlar , reaktif oksijen türleri ve reaktif azot türlerini içerir . Bu sekresyonlar, hava yolunu enfeksiyondan uzak tutmaya yardımcı olmak için doğrudan antimikrobiyal olarak işlev görebilir. Geleneksel bağışıklık hücrelerini ve diğerlerini enfeksiyon bölgesine alan çeşitli kemokinler ve sitokinler de salgılanır.
Sürfaktan bağışıklık fonksiyonu esas olarak iki proteine atfedilir: SP-A ve SP-D. Bu proteinler patojenlerin yüzeyindeki şekerlere bağlanabilir ve böylece fagositler tarafından alınması için opsonize edilebilir . Ayrıca enflamatuar yanıtları düzenler ve adaptif bağışıklık tepkisi ile etkileşime girer. Sürfaktan bozulması veya inaktivasyonu, akciğer iltihabı ve enfeksiyonuna karşı artan duyarlılığa katkıda bulunabilir. [31]
Solunum sisteminin çoğu, lenfositler gibi beyaz kan hücreleri üreten mukozaya bağlı lenfoid doku içeren mukoza zarlarıyla kaplıdır .
Alveolar çöküşün önlenmesi
Akciğerler , tip II alveoler hücreler tarafından oluşturulan bir yüzey aktif lipoprotein kompleksi (fosfolipoprotein) olan bir yüzey aktif madde oluşturur . Alveollerin içlerini çizen ve suyun yüzey gerilimini azaltan ince sulu tabakanın yüzeyinde yüzer.
Sulu bir yüzeyin (su-hava arayüzü) yüzey gerilimi bu yüzeyin küçülme eğilimindedir. [6] Bu yüzey akciğerlerin alveollerinde olduğu gibi kıvrıldığında, yüzeyin büzülmesi alveollerin çapını azaltır. Su-hava arayüzünün eğriliği ne kadar akut olursa , alveolusun çökme eğilimi o kadar büyük olur . [6] Bu üç etkileri vardır. İlk olarak alveollerin içindeki yüzey gerilimi, inhalasyon sırasında alveollerin genişlemesine direnir (yani akciğeri sertleştirir veya uyumlu değildir). Yüzey aktif madde yüzey gerilimini azaltır ve böylece akciğerleri daha uyumlu hale getirir, yoksa oradakinden daha az sert. İkinci olarak, alveollerin çapları solunum döngüsü sırasında artar ve azalır. Bu, alveollerin , soluma sonunda, soluma sonunda olan daha büyük bir çökme eğilimi (yani atelektazi neden olduğu ) anlamına gelir . Yüzey aktif madde sulu yüzeyde yüzdüğü için, ekshalasyon sırasında alveoller küçüldüğünde molekülleri daha sıkı bir şekilde paketlenir. [6]Bu, alveoller büyük olduklarından daha küçük olduğunda (yüzey aktif madde molekülleri daha geniş aralıklı olduğunda, inhalasyonun sonunda olduğu gibi) daha büyük bir yüzey gerilimi azaltıcı etkiye sahip olmalarına neden olur. Alveollerin çökme eğilimi, bu nedenle ekshalasyonun sonunda, inhalasyonun sonundaki ile hemen hemen aynıdır. Üçüncü olarak, alveolleri kaplayan kavisli sulu tabakanın yüzey gerilimi, akciğer dokularından alveollere su çekme eğilimindedir. Sürfaktan bu tehlikeyi ihmal edilebilir seviyelere düşürür ve alveolleri kuru tutar. [6] [32]
Sürfaktan üretemeyen erken dönem bebeklerin , her nefes aldıklarında çökme eğilimi gösteren akciğerleri vardır. Tedavi edilmediği sürece, solunum sıkıntısı sendromu olarak adlandırılan bu durum ölümcüldür. Tavuk akciğerlerinden alınan hücreler kullanılarak yapılan temel bilimsel deneyler, tip II alveoler hücrelerin gelişimini ilerletmek için steroid kullanma potansiyelini desteklemektedir . [33] Bir kez Aslında, erken doğum tehdit, nitelikli doğum geciktirmek için yapılır ve bir dizi steroid enjeksiyonu sıkça akciğer olgunlaşmasını teşvik etmek için, bu gecikme sırasında anneye uygulanır. [34]
Tüm vücut fonksiyonlarına katkılar
Akciğer damarları, embolizm ile pulmoner dolaşımına gelmiş olabilecek pıhtıları , genellikle bacaklardaki derin damarlardan çözen bir fibrinolitik sistem içerir . Ayrıca sistemik arteriyel kana giren çeşitli maddeleri salgılarlar ve pulmoner arter yoluyla onlara ulaşan sistemik venöz kandan diğer maddeleri çıkarırlar. Bazı prostaglandinler dolaşımdan çıkarılırken, diğerleri akciğerlerde sentezlenir ve akciğer dokusu gerildiğinde kana salınır.
Akciğerler bir hormonu aktive eder. Fizyolojik olarak inaktif dekapeptid anjiyotensin I , pulmoner dolaşımda aldosteron- salınan oktapeptide, anjiyotensin II’ye dönüştürülür . Reaksiyon diğer dokularda da görülür, ancak akciğerlerde özellikle belirgindir. Anjiyotensin II ayrıca arteriyolar duvarlar üzerinde doğrudan bir etkiye sahiptir , bu da arteriyolar vazokonstriksiyona ve dolayısıyla arteriyel kan basıncında bir artışa neden olur . [35] Bu aktivasyondan sorumlu büyük miktarlarda anjiyotensin dönüştürücü enzim endotelyal hücrelerin yüzeylerinde bulunur.alveoler kılcal damarların. Dönüştürücü enzim ayrıca bradikini inaktive eder . Alveolar kılcal damarlar arasındaki dolaşım süresi bir saniyeden azdır, ancak akciğerlere ulaşan anjiyotensin I’in% 70’i kılcal damarlardan tek bir seferde anjiyotensin II’ye dönüştürülür. Pulmoner endotel hücrelerinin yüzeyinde dört peptidaz daha tespit edilmiştir.
Seslendirme
Gazın gırtlak , farinks ve ağızdan hareketi, insanların konuşmasına veya fonatlamasına izin verir . Kuşlarda seslendirme veya şarkı söyleme, trakeanın tabanında bulunan bir organ olan syrinx yoluyla gerçekleşir . Larinks ( vokal kordlar ) boyunca, insanlarda ve syrinx’te, kuşlarda akan havanın titreşimi sese neden olur. Bu nedenle, gaz hareketi iletişim amaçları için hayati önem taşımaktadır .
Sıcaklık kontrolü
Panting köpek, kedi, kuş ve diğer hayvanlarda ağızdaki tükürüğün buharlaştırılması (yerine deride ter buharlaştırılması) ile, vücut sıcaklığının azaltılması için bir araç sağlamaktadır.
Klinik önemi
Solunum sistemi bozuklukları birkaç genel gruba ayrılabilir:
- Hava yolu obstrüktif koşulları (örn. Amfizem , bronşit , astım )
- Pulmoner kısıtlayıcı durumlar (örn. Fibroz , sarkoidoz , alveoler hasar, plevral efüzyon )
- Vasküler hastalıklar (örn., Pulmoner ödem , pulmoner emboli , pulmoner hipertansiyon )
- Bulaşıcı, çevresel ve diğer “hastalıklar” (örn. Pnömoni , tüberküloz , asbest , partikül kirleticiler )
- Primer kanserler (örn. Bronşiyal karsinom , mezotelyoma )
- İkincil kanserler (örneğin, vücudun başka yerlerinde ortaya çıkan, ancak kendilerini akciğerlerde tohumlayan kanserler)
- Yetersiz yüzey aktif madde (örn. Erken dönem bebeklerde solunum sıkıntısı sendromu ).
Solunum sistemi bozuklukları genellikle bir pulmonolog ve solunum terapisti tarafından tedavi edilir .
Nefes alamama veya nefes almada yetersizlik olduğunda tıbbi bir ventilatör kullanılabilir.
İstisnai memeliler
Atlar
Atlar zorunlu burun solunumlarıdır, yani ağızlarından nefes alma seçeneğine sahip olmadıkları ve burunlarından hava almaları gerektiği için diğer birçok memeliden farklı oldukları anlamına gelir.
Filler
Fil hiçbir olduğu bilinen tek memeli olduğunu plevral boşluk . Aksine, parietal ve viseral plevra hem yoğun bağ dokusundan oluşur hem de gevşek bağ dokusu yoluyla birbirine bağlanır. [36] Bu plevral boşluğun, alışılmadık derecede kalın bir diyaframla birlikte , şnorkel olarak ortaya çıkan gövdesinden nefes alarak filin uzun süre su altında kalmasına izin veren evrimsel uyarlamalar olduğu düşünülmektedir . [37]
Filde akciğerler diyaframa bağlanır ve nefes alma göğüs kafesinin genişlemesinden ziyade esas olarak diyaframa dayanır. [38]
Kuşlar

15 Kuşlarda hava keselerinin ve akciğerlerin düzenlenmesi

16 Kuşun solunum sisteminin anatomisi, trakea, primer ve pulmoner bronşlar, dorso- ve ventro-bronşların ikisi arasında çalışan parabronchi ile ilişkilerini gösterir. Posterior ve anterior hava keseleri de endikedir, ancak ölçeklenmez.

Şekil 17 Soluma sırasında göğsün hareketini gösteren bir güvercin iskeleti. Ok 1 , vertebral kaburgaların hareketini gösterir. Ok 2, sternumun (ve omurganın ) ardışık hareketini gösterir . İki hareket, kuş gövdesinin göğüs kısmının dikey ve enine çaplarını arttırır.
Anahtar:
1. kafatası ;
2. servikal vertebralar ;
3. furküla ;
4. korakoid ;
5. vertebral kaburgalar ;
6. sternum ve omurgası ;
7. patella ;
8. tarsus ;
9. basamak;
10. tibia ( tibiotarsus );
11. fibula ( tibiotarsus );
12. uyluk kemiği ;
13. ischium ( innominat );
14. pubis (innominate);
15. ilium (innominat);
16. kuyruk omurları ;
17. pigostyle ;
18. synsacrum ;
19. skapula ;
20. dorsal omurlar ;
21. humerus ;
22. ulna ;
23. yarıçap ;
24. bilek ( carpometacarpus );
25. metacarpus (carpometacarpus );
26. basamak ;
27. alula

18 Kuşlarda soluma-ekshalasyon döngüsü.
Kuş akciğerleri, karşılaştırılabilir büyüklükteki memelilerden daha küçüktür, ancak hava keseleri , memelilerde körük görevi gören alveollere adanmış% 7’ye kıyasla, toplam vücut hacminin% 15’ini oluşturur . [40]
Soluma ve ekshalasyon, hem torako-abdominal boşluğun (veya coelom ) hem abdominal hem de kostal kaslarını kullanarak dönüşümlü olarak arttırılarak ve azaltılarak gerçekleştirilir . [41] [42] [43] İnhalasyon sırasında vertebral kaburgalara bağlı kaslar (Şek. 17) onları öne ve dışa doğru eğirir. Bu, sternumu (belirgin omurgasıyla ) aynı yönde alarak neredeyse dik açılarda, aşağı ve ileri doğru tutturuldukları sternal kaburgaları iter (Şekil 17). Bu, gövdenin torasik kısmının hem dikey hem de enine çaplarını arttırır. Özellikle posteriorun ileri ve aşağı hareketisternumun ucu karın duvarını aşağı doğru çekerek gövdenin o bölgesinin hacmini arttırır. [41] Tüm gövde boşluğunun hacmindeki artış tüm torako-abdominal hava keselerindeki hava basıncını düşürerek, aşağıda açıklandığı gibi hava ile doldurmalarına neden olur.
Ekshalasyon sırasında, sternum ve vertebral kaburgalara ön tarafa ve pelvise (Şekil 17’deki pubis ve ilium) posterior (karın duvarının bir kısmını oluşturan ) bağlanan dış oblik kas , karın içeriğini sıkıştırarak inhalasyon hareketini tersine çevirir, böylece tüm hava keselerindeki basıncı arttırır. Bu nedenle hava, ekshalasyon eyleminde solunum sisteminden atılır. [41]

Şekil 19 Kuşların akciğerlerindeki çapraz akım solunum gazı değiştirici. Hava, hava keselerinden parabronchi boyunca tek yönlü olarak (şemada sağdan sola) zorlanır. Pulmoner kılcal damarlar parabronchi’yi gösterilen şekilde çevreler (parabronşusun altından şemada üzerine akan kan). [41] [44] Oksijen içeriği yüksek olan kan veya hava kırmızıyla gösterilir; oksijen açısından fakir hava veya kan mor-mavinin çeşitli tonlarında gösterilmiştir.
İnhalasyon sırasında hava girer nefes borusunu burun delikleri ve ağız yoluyla ve devam sadece ötesinde syrinx ikiye trakea dalları bu noktada birincil bronşlarda iki akciğerler (Şek. 16) gidiş. Birincil bronşlar, ventrobronchi adı verilen bir dizi paralel dal ve biraz da eşdeğer bir dorsobronchi seti veren intrapulmoner bronşlar olmak için akciğerlere girer (Şekil 16). [41] İntrapulmoner bronşların uçları havayı kuşun kuyruk ucundaki arka hava keselerine boşaltır . Dorsolomber ventrobronchi her bir çifti bir büyük paralel mikroskopik hava kılcal sayısına (veya ile bağlanır parabronchi ) gaz değişimioluşur (Şekil 16). [41] kuş bir nefes çektiğinde, trakeal hava, arka hava keseleri içerisine intrapulmoner bronş akar olarak içine dorso değil içine, bronşlar ventro bronşlar (Şek. 18). Bu, solunan havayı ventrobronchi’nin açıklıklarından uzağa, intrapulmoner bronşun dorsobronchi ve arka hava keselerine doğru devam ettirmesine yönlendiren bronşiyal mimariden kaynaklanmaktadır. [45] [46] [47] Dorsobronchi’den solunan hava parabronchi’den (ve dolayısıyla gaz eşanjöründen) havanın sadece genişleyen ön hava keselerine kaçabileceği ventrobronchi’ye akar. Böylece, inhalasyon sırasında hem posterior hem de ön hava keseleri genişler,[41] arka hava keseleri taze solunan hava ile doldurulurken, ön hava keseleri akciğerlerden yeni geçen “kullanılmış” (oksijen fakiri) hava ile doldurulur.
Ekshalasyon sırasında, posterior hava keselerindeki (inhalasyon sırasında temiz hava ile doldurulmuş) basınç, yukarıda tarif edilen eğik kasın kasılması nedeniyle artar. Arka hava keselerinden dorsobronchi ve intrapulmoner bronşlara kadar birbirine bağlı açıklıkların aerodinamiği, havanın intrapulmoner bronşları geri döndürmek yerine akciğerlerden (dorsobronchi yoluyla) ayrılmasını sağlar (Şekil 18). [45] [47]Dorsobronchi’den arka hava keselerinden gelen taze hava parabronchi’den (inhalasyon sırasında meydana gelenle aynı yönde) ventrobronchi’ye akar. Ventrobronchi ve ön hava keselerini intrapulmoner bronşlara bağlayan hava geçitleri, bu iki organdan “harcanan”, oksijen yetersiz havayı dışarıya kaçtığı trakeaya yönlendirir. [41] Oksijenli hava bu nedenle parabronchi boyunca tek bir doğrultuda (tüm solunum döngüsü sırasında) sürekli olarak akar. [48]
Kuş akciğeri boyunca kan akışı, parabronchi’den hava akışına dik açılardadır ve çapraz akım akış değişim sistemi oluşturur (Şekil 19). [39] [41] , [44] , oksijenin kısmi basıncının parabronchi O olarak, uzunlukları boyunca azalır 2 kana yayıldığı. Eşanjörü hava akışının girişine yakın bir yerde bırakan kan kılcal damarları , parabronchi’nin çıkış ucunun yakınında kalan kılcal damarlardan daha fazla O 2 alır . Tüm kılcal damarların içeriği karıştığında, karışık pulmoner venöz kanın nihai kısmi oksijen basıncı, solunan havanınkinden daha yüksektir, [41] [44], ancak yine de solunan havanın yarısından daha azdır, [41]böylece memelilerin körük tipi ciğerlerinde olduğu gibi kabaca aynı sistemik arteriyel kan kısmi oksijen basıncına ulaşılması . [41]
Trakea ölü bir boşluk alanıdır : ekshalasyonun sonunda içerdiği oksijen bakımından fakir hava, posterior hava keselerine ve akciğerlerine tekrar giren ilk havadır. Memeli solunum sistemine kıyasla, bir kuştaki ölü boşluk hacmi, aynı büyüklükteki memelilerden ortalama 4.5 kat daha fazladır. [40] [41] Uzun boyunlu kuşların kaçınılmaz olarak uzun trakeaları olacaktır ve bu nedenle memelilerin daha büyük ölü boşluk hacimlerine izin vermek için yaptıkları nefeslerden daha derin nefes almaları gerekir. Bazı kuşlarda (örneğin ötücü kuğu , Cygnus cygnus , beyaz kaşıkçı , Platalea leucorodia , boğmaca vinç ,Grus americana ve kasklı curassow , Pauxi pauxi ) bazı vinçlerin 1,5 m uzunluğunda olabileceği trakea, [41] vücut içinde ileri geri sarılır ve ölü boşluk havalandırmasını büyük ölçüde arttırır. [41] Bu olağanüstü özelliğin amacı bilinmiyor.
Sürüngenler
Amfibi
Hem akciğerler hem de deri , amfibilerde solunum organları olarak hizmet eder . Amfibilerde akciğerlerin havalandırılması pozitif basınçlı ventilasyona dayanır . Kaslar ağız boşluğunun tabanını indirir, genişletir ve burun deliklerinden ağız boşluğuna hava çeker . Burun delikleri ve ağız kapalıyken, ağız boşluğunun tabanı yukarı itilir, bu da havayı trakeadan akciğerlere doğru zorlar. Bu hayvanların derisi yüksek oranda vaskülarize ve nemli olup , özel hücrelerden mukus salgılanmasıyla nem korunur ve kutanöz solunumda rol oynar. Akciğerler kan ve çevresel hava (su dışındayken) arasında gaz alışverişi için birincil organlara sahipken, cildin benzersiz özellikleri, amfibiler oksijen açısından zengin suya batırıldığında hızlı gaz değişimine yardımcı olur. [50] Bazı amfibiler kendi gelişiminin erken evrelerinde (örn ya, solungaçları vardır tetarlar ait kurbağalar diğerleri (örneğin bazı yetişkinlikte bunları muhafaza ederken,) semender ). [39]
Balık

Şek. 21. kapakçık ya da bir solungaç kapak turna ortaya çıkarmak için açık çekilmesinden solungaç kemeri taşıyan filamanlar .

Şekil 22. Bir eş-akım ve bir ters-akım akış değişim sisteminin operasyonları ve etkileri arasındaki bir karşılaştırma, sırasıyla üst ve alt diyagramlar ile tasvir edilmektedir. Her ikisinde de kırmızının maviden daha yüksek bir değere (örneğin sıcaklık veya bir gazın kısmi basıncı) sahip olduğu ve kanallarda taşınan özelliğin kırmızıdan maviye aktığı varsayılmaktadır. Balıklarda, solungaçlardaki kan ve suyun ters akım (alt diyagramı) ortamdan oksijen çıkarmak için kullanılır. [51] [52] [53]
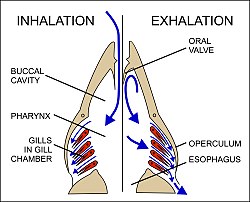
23 Kemikli balıklarda solunum mekanizması. Solunum süreci solda, ekshalasyon süreci sağdadır. Suyun hareketi mavi oklarla gösterilir.
Oksijen suda az çözünür. Tam gazlı tatlı su nedenle içeren 8-10 ml o 2 / litre O ile karşılaştırıldığında 2 210 ml / deniz seviyesinde hava litre konsantrasyonda. [54] Ayrıca, difüzyon katsayısı (yani, kuru olan standart koşullar altında, düşük konsantrasyonda bir yüksek konsantrasyonlu bir bölgesinden bir madde dağılır) solunum gazlarının ise , tipik olarak 10,000 hızlı hava içinde suya göre . [54] Bu nedenle, oksijen, örneğin, 17.6 mm ‘lik bir difüzyon katsayısı 2 havada / s, ama sadece 0.0021 mm 2 / su içinde s. [55] [56] [57][58] , karbon dioksit için karşılık gelen değerler, 16 mm 2 havada / s ve 0.0016 mm 2 / su içinde s. [57] [58] Bu, bir gaz eşanjörü ile temas halindeki sudan oksijen alındığında, oksijen bakımından zengin bölgelerden, oksijen değiştirici bölgelerinden meydana gelenlerden daha uzak mesafelerden gelen oksijen ile çok daha yavaş bir şekilde değiştirildiği anlamına gelir. Balık bu problemlerle ilgilenen solungaçlar geliştirmiştir. Solungaçlar, lamellere daha da bölünen filaman içeren özel organlardır. Lameller yoğun bir ince duvarlı kılcal ağ içerirbüyük bir gaz değişim yüzey alanını, üzerlerinden geçen çok büyük miktarlarda suya maruz bırakır. [59]
Solungaçlar , sudan oksijen alımının verimliliğini artıran bir ters akım değişim sistemi kullanır. [51] [52] [53] Ağızdan alınan taze oksijenli su, solungaçlardan bir yönde kesintisiz olarak “pompalanır”, lameldeki kan ters yönde akar, ters akan kan ve su akışını oluşturur (Şek. 22), balığın hayatta kalmasına bağlıdır. [53]
Su, operculum (solungaç kapağı) kapatılarak ve ağız boşluğu büyütülerek ağızdan içeri çekilir (Şek. 23). Eşzamanlı olarak solungaç odaları genişler, orada ağızdan daha düşük bir basınç üreterek su solungaçların üzerinden akmasına neden olur. [53] , ağız boşluğu, böylece ağız (Şek. 23), suyun geri akışını önleyen, pasif ağız valflerin kapanmasına neden olan daha sonra sözleşmeler. [53] [60] Bunun yerine, ağızdaki su solungaçların üzerine zorlanırken, solungaç odaları, içerdikleri suyu operküler açıklıklardan boşaltmak için büzülür (Şek. 23). İnhalasyon fazı sırasında solungaç odasına geri akış, ventroposterior boyunca bir zar tarafından önleniroperculum sınırı (Şekil 23’te soldaki diyagram). Böylece ağız boşluğu ve solungaç odaları, solungaçların bir yönde sabit bir su akışını sağlamak için dönüşümlü olarak emme pompası ve basınç pompası görevi görür. [53] Lamel kılcal damarlardaki kan suyun akış yönünün tersi yönde aktığı için, bunun sonucunda kan ve suyun ters akım akışı, her bir kılcal damarın tüm uzunluğu boyunca oksijen ve karbondioksit için dik konsantrasyon gradyanlarını korur (Şek. 22). Dolayısıyla oksijen, gradyanını sürekli olarak kana ve karbon dioksiti de gradyanını suya doğru dağıtabilir. [52]Ters akım değişim sistemleri teorik olarak, bir solunum gazının eşanjörün bir tarafından diğerine neredeyse tamamen aktarılmasına izin verse de, solungaçlar üzerinden akan suda oksijenin% 80’inden azı genellikle kana aktarılır. [51]
Bazı aktif pelajik köpekbalıklarında, su hareket halindeyken “koç havalandırma” olarak bilinen bir işlemle ağızdan ve solungaçların üzerinden geçer. [61] dinlenmede iken, en köpekbalıkları en kemikli balıklar gibi, bu oksijenli su onların solungaçları üzerinde akmaya devam sağlamak için, onların solungaçları üzerinde su pompası. Ancak az sayıda tür solungaçlarından su pompalama yeteneğini kaybetmiştir ve dinlenmeden yüzmelidir. Bu türler zorunlu koç vantilatörleridir ve hareket edemezlerse muhtemelen boğulurlar . Koç havalandırmasının zorunlu olması bazı pelajik kemikli balık türleri için de geçerlidir. [62]
Su yüzeyinin üstünden yutulan havadan kısa süreliğine oksijen alabilen birkaç balık vardır. Böylece Akciğer Balığı bir veya iki akciğere sahiptir ve labirent balıkları , bu balık alt sınırını karakterize eden özel bir “labirent organı” geliştirmiştir. Labirent organ, çok katlı bir supra branşiyal aksesuar solunum organıdır . İlk solungaç kemerinin epibranşiyal kemiğinin vaskülarize bir genişlemesi ile oluşur ve havada solunum için kullanılır . [63]
Bu organ, labirent balıklarının solungaçları kullanarak içinde bulundukları sudan almak yerine doğrudan havadan oksijen almasına izin verir . Labirent organ, solunan havadaki oksijenin kan dolaşımına emilmesine yardımcı olur . Sonuç olarak, labirent balıkları, nemli kalmaları koşuluyla, etraflarındaki havayı soluyabildikleri için sudan kısa bir süre hayatta kalabilirler.
Labirent balığı fonksiyonel labirent organlarıyla doğmaz. Organın gelişimi kademelidir ve çoğu çocuk labirent balığı tamamen solungaçlarıyla nefes alır ve büyüdüklerinde labirent organlarını geliştirir. [63]
omurgasızlar
Eklembacaklılar
Bazı yengeç türleri, braniostegal akciğer adı verilen bir solunum organı kullanır . [64] Bu solungaç benzeri yapısı daha sudan daha havadan oksijen alma için uygun gaz değişimi için yüzey alanını arttırır. En küçük örümcek ve akarlardan bazıları , vücudun yüzeyinden gaz alışverişi yaparak nefes alabilir. Daha büyük örümcekler, akrepler ve diğer eklembacaklılar ilkel bir kitap akciğeri kullanır .
Haşarat
En nefes pasif olarak yoluyla böcek spiracles (özel açıklıklar dış iskelet ) ve hava ‘trachaea’ çapları nispeten büyük olduğunda adı verilen daha küçük ve daha küçük bir seri tüp vasıtasıyla vücudun her yerinde ulaşır ve ‘ tracheoles ‘ onların çapları çok küçük. Trakeoller vücuttaki bireysel hücrelerle temas eder. [39] Kısmen kaslar gibi dokular aktif olduğunda ve oksijen için yüksek bir talebe sahip olduklarında havayı aktif hücrelere yakınlaştırarak bireysel trakeollerden çekilebilen sıvı ile doldurulurlar. [39]Bu muhtemelen aktif kaslarda bir ozmotik gradyana neden olan, suyu trakeollerden ve aktif hücrelere hareket ettiren laktik asit birikimi ile ortaya çıkar. Gazların difüzyonu küçük mesafelerde etkilidir, ancak daha büyük mesafelerde değil, böceklerin nispeten küçük olmasının nedenlerinden biridir. Bazı Collembola gibi spiracles ve trachaea olmayan böcekler, derilerinden doğrudan gazların yayılmasıyla nefes alırlar. [65]
Bir böceğin sahip olduğu spiracles sayısı türler arasında değişkendir, ancak her zaman çift olarak, vücudun her iki tarafında bir tane ve genellikle segment başına bir çift olarak gelirler. Diplura’nın bazıları on bir, toraks üzerinde dört çift ile var, ancak Yusufçuklar ve Çekirge gibi eski böcek formlarının çoğunda iki torasik ve sekiz karın spiracle var. Bununla birlikte, geri kalan böceklerin çoğunda daha az vardır. Solunum için hücrelere oksijen iletilen trakeoller seviyesindedir.
Böceklerin bir zamanlar , gazların trakeal sisteme basit difüzyonu ile sürekli olarak çevre ile gaz değiştirdiklerine inanılıyordu . Bununla birlikte, daha yakın zamanlarda, böcek ventilatör paternlerinde büyük farklılıklar belgelenmiştir ve böcek solunumu oldukça değişken görünmektedir. Bazı küçük böcekler sürekli solunum hareketleri göstermez ve spiracles kas kontrolünden yoksun olabilir. Diğerleri ise, kullanan kas kasılması ve karın koordine spiracle daralma ve çevrimsel gaz değiştirme kalıplarını üretmek için ve atmosfere su kaybını azaltmak için gevşeme ile birlikte. Bu modellerin en uç şekli süreksiz gaz değişim çevrimleri olarak adlandırılır . [66]
Yumuşakçalar
Yumuşakçalar genellikle sulu ortam ile bunların dolaşım sistemleri arasında gaz alışverişine izin veren solungaçlara sahiptir. Bu hayvanlar ayrıca oksijen yakalayan molekülü olarak hemosiyanin içeren kanı pompalayan bir kalbe sahiptir . [39] Bu nedenle, bu solunum sistemi omurgalı balık benzerdir. Gastropoda solunum sistemi ya solungaçlar veya akciğer içerebilir.
Bitkiler
Bitkiler fotosentez sürecinde karbondioksit gazı kullanır ve atık olarak oksijen gazını verir. Fotosentez kimyasal denklem 6 CO 2 (karbon dioksit) ve 6H 2 güneş ışığının mevcudiyetinde Cı yapar O (su), 6 H 12 O 6 (glikoz) ve 6 O 2 (oksijen). Fotosentez, güneş ışığından elde edilen enerji deposu olarak karbon atomları üzerindeki elektronları kullanır. [67] Solunum fotosentez tersidir. Hücrelerdeki kimyasal reaksiyonlara güç vermek için enerjiyi geri kazanır. Bunu yaparken karbon atomları ve elektronları oksijen oluşturan CO ile birleştirilir.2 hem hücrelerden hem de organizmadan kolayca uzaklaştırılabilir. Bitkiler enerjiyi yakalamak için her iki işlemi, fotosentezi ve kullanmak için oksidatif metabolizmayı kullanırlar.
Bitki solunumu difüzyon süreci ile sınırlıdır . Bitkiler , yapraklarının altlarında ve bazen anatomilerinin diğer kısımlarında açılabilen ve kapanabilen stomalar olarak bilinen deliklerden karbondioksit alırlar . Çoğu bitki katabolik süreçler için biraz oksijene ihtiyaç duyar (enerjiyi serbest bırakan parçalanma reaksiyonları). Ancak saatte kullanılan O 2 miktarı, yüksek oranda aerobik metabolizma gerektiren aktivitelerde yer almadığından azdır . Bunlar CO ihtiyaç olarak hava için gereksinim, ancak çok yüksek olduğu 2 çevre hava sadece% 0.04 teşkil fotosentez için. Bu nedenle, 1 g glikoz yapmak için tüm CO2 ile ilgili en az bir deniz seviyesinde hava 18.7 litre. Ancak fotosentetik işlemdeki verimsizlikler çok daha fazla miktarda havanın kullanılmasına neden olur. [67] [68]
Kaynakça
- ^ Campbell, Neil A. (1990). Biyoloji (2. baskı). Redwood City, Kaliforniya: Benjamin / Cummings Pub. Co. s. 834-835. ISBN 0-8053-1800-3.
- ^ Hsia, CC; Hyde, DM; Weibel, ER (15 Mart 2016). “Akciğer Yapısı ve Gaz Değişiminin Temel Zorlukları” . Kapsamlı Fizyoloji . 6 (2): 827-95. doi : 10.1002 / cphy.c150028 . PMC 5026132 . PMID 27065169 .
- ^ West, John B. (1995). Solunum fizyolojisi – temeller . Baltimore: Williams ve Wilkins. s. 1-10 . ISBN 0-683-08937-4.
- ^ Yukarıya atlayın:a b Gilroy, Anne M .; MacPherson, Brian R .; Ross, Lawrence M. (2008). Anatomi Atlası . Stuttgart: Thieme. ss. 108–111. ISBN 978-1-60406-062-1.
- ^ Yukarıya atlayın:a b Pocock, Gillian; Richards, Christopher D. (2006). İnsan fizyolojisi: tıbbın temeli (3. baskı). Oxford: Oxford Üniversitesi Yayınları. s. 315-317. ISBN 978-0-19-856878-0.
- ^ Aşağıya atlayın:a b c d e f g h i j k l m n o p q r s t u v Tortora, Gerard J .; Anagnostakos, Nicholas P. (1987). Anatomi ve fizyoloji prensipleri (Beşinci Baskı). New York: Harper & Row, Yayıncılar. sayfa 556-586 . ISBN 0-06-350729-3.
- ^ Kacmarek, Robert M .; Dimas, Steven; Mack, Craig W. (13 Ağustos 2013). Solunum Bakımının Temelleri – E-Kitap . Elsevier Sağlık Bilimleri. ISBN 9780323277785.
- ^ Netter, Frank H. (2014). Öğrenci Dahil İnsan Anatomisi Atlası İnteraktif Yardımcılar ve Kılavuzlar’a danışın (6. baskı). Philadelphia, Penn .: WB Saunders Co. p. 200. ISBN 978-1-4557-0418-7.
- ^ Maton, Anthea; Jean Hopkins; Charles William McLaughlin; Susan Johnson; Maryanna Quon Warner; David LaHart; Jill D.Wright (1993). İnsan Biyolojisi ve Sağlık . wood Cliffs, New Jersey, ABD: Prentice Salonu. ISBN 0-13-981176-1.[ sayfa gerekli ]
- ^ Yukarıçık :a b c Fowler WS (1948). “Akciğer Fonksiyon çalışmaları. II. Solunum ölü boşluğu”. Am. J. Physiol . 154 (3): 405-416. doi : 10.1152 / ajplegacy.1948.154.3.405 . PMID 18101134 .
- ^ “anatomik ölü boşluk” . TheFreeDictionary.com .
- ^ Yukarıçık :a b Tortora, Gerard J .; Anagnostakos, Nicholas P. (1987). Anatomi ve fizyoloji prensipleri (Beşinci Baskı). New York: Harper & Row, Yayıncılar. sayfa 570-572 . ISBN 0-06-350729-3.
- ^ Yukarıya atlayın:a b Koen, Chrisvan L .; Koeslag, Johan H. (1995). “Subatmosferik intraplevral ve intrakraniyal basınçların stabilitesi üzerine”. Fizyolojik Bilimlerde Haberler . 10 (4): 176-178. doi : 10.1152 / fizyologyonline . 1995.10.4.176 .
- ^ Yukarıya atlayın:a b West, JB (1985). Solunum fizyolojisi: temeller . Baltimore: Williams ve Wilkins. s. 21–30, 84–84, 98-101.
- ^ Burke, TV; Küng, M; Burki, NK (1989). “Astımlı hastalarda histamin kaynaklı bronkokonstriksiyon sırasında pulmoner gaz değişimi” . Göğüs . 96 (4): 752-6. doi : 10.1378 / göğüs.9.4.752 . PMID 2791669 .
- ^ Taylor, D (1996). “Valsalva Manevrası: Eleştirel bir inceleme” . Güney Pasifik Sualtı Tıbbı Derneği Dergisi . 26 (1). ISSN 0813-1988 . OCLC 16986801 . Erişim tarihi: 14 Mart 2016 .
- ^ Maton, Anthea; Hopkins, Jean Susan; Johnson, Charles William; McLaughlin, Maryanna Quon; Warner, David; LaHart Wright, Jill (2010). İnsan Biyolojisi ve Sağlık . Englewood Kayalıkları: Prentice Hall. ss. 108–118. ISBN 978-0134234359.
- ^ En üste atlayın:a b c Williams, Peter L .; Warwick, Roger; Dyson, Mary; Bannister, Lawrence H. (1989). Gray’in Anatomisi (Otuz yedinci baskı). Edinburgh: Churchill Livingstone. sayfa 1278-1282. ISBN 0443-041776.
- ^ Lovelock, James (1991). Şifa Gaia: Gezegen için pratik tıp . New York: Uyum Kitapları. 21-23, 73-88. ISBN 0-517-57848-4.
- ^ Shu, BC; Chang, YY; Lee, FY; Tzeng, DS; Lin, HY; Akciğer, FW (2007-10-31). “Hiperventilasyon sendromu olan genç erkeklerde ebeveyn bağlanması, premorbid kişilik ve ruh sağlığı”. Psikiyatri Araştırmaları . 153 (2): 163-70. doi : 10.1016 / j.psychres.2006.05.006 . PMID 17659783 .
- ^ Henry RP, Swenson ER (Haziran 2000). “Omurgalı gaz değişim organlarında karbonik anhidrazın dağılımı ve fizyolojik önemi”. Solunum Fizyolojisi . 121 (1): 1–12. doi : 10.1016 / S0034-5687 (00) 00110-9 . PMID 10854618 .
- ^ Diem, K .; Lentner, C. (1970). “Kan – İnorganik maddeler”. in: Bilimsel Tablolar (Yedinci Baskı). Basle, İsviçre: CIBA-GEIGY Ltd. s. 571.
- ^ Yukarı atlayın:a b “Solunum” . Harvey Projesi . Erişim tarihi: 27 Temmuz 2012 .
- ^ En üste atla:a b “Online yüksek irtifa oksijen hesaplayıcısı” . altitude.org. Arşivlenmiş orijinal 29 Temmuz 2012 tarihinde . Erişim tarihi: 15 Ağustos 2007 .
- ^ Tyson, PD; Preston-White, RA (2013). Hava ve iklim Güney Afrika . Cape Town: Oxford Üniversitesi Yayınları. s. 3-10, 14–16, 360. ISBN 9780195718065.
- ^ Diem, K .; Lenter, C. (1970). Bilimsel Tablolar (Yedinci Baskı). Basle, İsviçre: Ciba-Geigy. sayfa 257-258.
- ^ Von Euler, ABD; Liljestrand, G. (1946). “Kedide pulmoner arteriyel kan basıncı üzerine gözlemler”. Açta Physiologica Scandinavica . 12 (4): 301–320. doi : 10.1111 / j.1748-1716.1946.tb00389.x .
- ^ “EPO Tespiti” . Dünya Dopingle Mücadele Ajansı . Erişim tarihi: 7 Eylül 2017 .
- ^ Yukarıçık :a b Tortora, Gerard J .; Anagnostakos, Nicholas P. (1987). Anatomi ve fizyoloji prensipleri (Beşinci Baskı). New York: Harper & Row, Yayıncılar. sayfa 444-445 . ISBN 0-06-350729-3.
- ^ Fisher JW, Koury S, Ducey T, Mendel S (1996). “Hipoksik maymun böbreklerinin interstisyel hücreleri tarafından eritropoietin üretimi”. İngiliz Hematoloji Dergisi . 95 (1): 27–32. doi : 10.1046 / j.1365-2141.1996.d01-1864.x . PMID 8857934 .
- ^ Wright, Jo Rae (2004). “Akciğer Sürfaktanının Konak Savunma Fonksiyonları”. Yenidoğanın Biyolojisi . 85 (4): 326-32’de açıklanmaktadır. doi : 10.1159 / 000078172 . PMID 15211087 .
- ^ West, John B. (1994). Solunum fizyolojisi – temeller . Baltimore: Williams ve Wilkins. s. 21–30, 84–84, 98-101 . ISBN 0-683-08937-4.
- ^ Sullivan, LC; Orgeig, S (2001). “Deksametazon ve epinefrin, embriyonik tavukların tip II hücrelerinde yüzey aktif madde salgısını uyarır”. Amerikan Fizyoloji Dergisi. Düzenleyici, Bütünleştirici ve Karşılaştırmalı Fizyoloji . 281 (3): R770-7. doi : 10.1152 / ajpregu.2001.281.3.r770 . PMID 11506991 .
- ^ Prematüre Bebekler, Akciğer Gelişimi ve Solunum Sıkıntısı Sendromu . Pregnancy-facts.com.
- ^ Kanaid, Hideo; Ichiki, Toshihiro; Nishimura, Junji; Hirano, Katsuya (2003-11-28). “Anjiyotensin II’nin Neden Olduğu Vazokonstriksiyonun Hücresel Mekanizması Belirlenmeye Devam Ediyor”. Dolaşım Araştırması . 93 (11): 1015-1017. doi : 10.1161 / 01.RES.0000105920.33926.60 . ISSN 0009-7330 . PMID 14645130 .
- ^ Batı, John B .; Ravichandran (1993). “Filde şnorkel nefes almak onun plevrasının eşsiz anatomisini açıklıyor”. Solunum Fizyolojisi . 126 (1): 1-8. doi : 10.1016 / S0034-5687 (01) 00203-1 . PMID 11311306 .
- ^ West, John B. (2002). “Filin neden plevral bir alanı yok?” . Haberler Physiol Sci . 17 (2): 47–50. doi : 10.1152 / nips.01374.2001 . PMID 11909991 .
- ^ Shoshani, Jeheskel (Aralık 1998). “Prozosid evrimi anlama: müthiş bir görev”. Ekoloji ve Evrimde Eğilimler . 13 (12): 480-487. doi : 10.1016 / S0169-5347 (98) 01491-8 . PMID 21238404 .
- ^ Yukarı atlayın:a b c d e f g Campbell, Neil A. (1990). Biyoloji (2. baskı). Redwood City, Kaliforniya: Benjamin / Cummings Pub. Co. s. 836-844. ISBN 0-8053-1800-3.
- ^ En üste atlayın:a b Whittow, G. Causey (2000). Sturkie’nin Kuş Fizyolojisi . San Diego, California: Akademik Basın. sayfa 233-241. ISBN 978-0-12-747605-6.
- ^ Kadar geçin:Bir b c d e f g h i j k l m n o Ritchson G. “BIO 554/754 – Böcekler: Kanatlı solunum” . Biyolojik Bilimler Bölümü, Eastern Kentucky Üniversitesi . Erişim tarihi: 2009-04-23 .
- ^ Storer, Tracy I .; Usinger, RL; Stebbins, Robert C .; Nybakken, James W. (1997). Genel Zooloji (altıncı baskı). New York: McGraw-Hill. sayfa 752-753 . ISBN 0-07-061780-5.
- ^ Romer, Alfred Sherwood (1970). Omurgalı cismi (Dördüncü baskı). Philadelphia: WB Saunders. sayfa 323-324 . ISBN 0-7216-7667-7.
- ^ En üste atlayın:a b c Scott, Graham R. (2011). “Yorum: Yüksek performans: yüksek irtifalarda uçan kuşların eşsiz fizyolojisi”. Deneysel Biyoloji Dergisi . 214 (Pt 15): 2455-2462. doi : 10.1242 / jeb.052548 . PMID 21753038 .
- ^ Yukarı atlayın:a b Maina, John N. (2005). Kuşların akciğer hava kesesi sistemi gelişimi, yapısı ve işlevi; 6 tablo ile . Berlin: Springer. 3.2. 3.3.3 “Akciğer”, “Havayolu (Bronchiol) Sistemi” 66-82. ISBN 978-3-540-25595-6.
- ^ Krautwald-Junghanns, Maria-Elisabeth; vd. (2010). Egzotik Evcil Hayvanların Tanısal Görüntüleme: Kuşlar, Küçük Memeliler, Sürüngenler . Almanya: Manson Yayınları. ISBN 978-3-89993-049-8.
- ^ Yukarıçık :a b Sturkie, PD (1976). Kuş Fizyolojisi . New York: Springer Verlag. s. 201. doi : 10.1007 / 978-1-4612-4862-0 . ISBN 978-1-4612-9335-4.
- ^ Ritchison, Gary. “Ornitoloji (Bio 554/754): Kuş Solunum Sistemi”. Doğu Kentucky Üniversitesi. Erişim tarihi: 2007-06-27.
- ^ Solunum sistemi . Britannica Ansiklopedisi.
- ^ Gottlieb, G; Jackson DC (1976). “Boğa kurbağasında solunum kontrolünde pulmoner ventilasyonun önemi”. J Physiol . 230 (3): 608-13. doi : 10.1152 / ajplegacy.1976.230.3.608 . PMID 4976 .
- ^ En üste atlayın:a b c Campbell, Neil A. (1990). Biyoloji (İkinci baskı). Redwood City, California: Benjamin / Cummings Yayıncılık Şirketi, ss. 836-838. ISBN 0-8053-1800-3.
- ^ En üste atlayın:a b c Hughes GM (1972). “Balık solungaçlarının morfometri”. Solunum Fizyolojisi . 14(1–2): 1–25. doi : 10.1016 / 0034-5687 (72) 90014-x . PMID 5042155 .
- ^ Yukarı atlayın:a b c d e f Storer, Tracy I .; Usinger, RL; Stebbins, Robert C .; Nybakken, James W. (1997). Genel Zooloji (altıncı baskı). New York: McGraw-Hill. sayfa 668-670 . ISBN 0-07-061780-5.
- ^ Yukarıçık :a b M. bv Roberts; Michael Reiss; Grace Monger (2000). İleri Biyoloji . Londra, İngiltere: Nelson. s. 164-165.
- ^ Cussler, EL (1997). Difüzyon: Akışkan Sistemlerinde Kütle Transferi (2. baskı). New York: Cambridge Üniversitesi Yayınları. ISBN 0-521-45078-0.
- ^ Welty, James R .; Wicks, Charles E .; Wilson, Robert E .; Rorrer, Gregory (2001). Momentum, Isı ve Kütle Transferinin Temelleri . Wiley. ISBN 978-0-470-12868-8.
- ^ Yukarıya atlayın:a b CRC Press Online: CRC Kimya ve Fizik El Kitabı, Bölüm 6, 91. Baskı
- ^ Yukarıya atla:a b Difüzyon
- ^ Newstead James D (1967) ‘de açıklanmaktadır. “Teleost solungaçların solunum lamellerinin ince yapısı”. Hücre ve Doku Araştırmaları . 79 (3): 396-428. doi : 10.1007 / bf00335484 . PMID 5598734 .
- ^ Romer, Alfred Sherwood ; Parsons, Thomas S. (1977). Omurgalı Vücut . Philadelphia, PA: Holt-Saunders Uluslararası. s. 316-327. ISBN 0-03-910284-X.
- ^ Gilbertson, Lance (1999). Zooloji Laboratuvarı El Kitabı . New York: McGraw-Hill. ISBN 0-07-237716-X.
- ^ William J. Bennetta (1996). “Derin Nefes” . Erişim tarihi: 2007-08-28 .
- ^ Yukarıya atlayın:a b Pinter, H. (1986). Labirent balığı. Barron Eğitim Serisi, Inc., ISBN 0-8120-5635-3
- ^ Halperin J, Ansaldo M, Pellerano GN, Luquet CM (Temmuz 2000). “Estuarin yengeç Chaimmagnathus granulatus Dana 1851 bimodal nefes – fizyolojik ve morfolojik çalışmalar”. Karşılaştırmalı Biyokimya ve Fizyoloji. Bölüm A, Moleküler ve Bütünleştirici Fizyoloji . 126(3): 341–9. doi : 10.1016 / S1095-6433 (00) 00216-6 . PMID 10964029 .
- ^ Dünya Yaşam Ağı, Böcek Morfolojisi ve Anatomisi . Earthlife.net. Erişim tarihi: 2013-04-21.
- ^ Lighton, JRB (Ocak 1996). “Böceklerde süreksiz gaz değişimi”. Annu Rev Entomol . 41 : 309-324’te açıklanmaktadır. doi : 10.1146 / annurev.en.41.010196.001521 . PMID 8546448 .
- ^ Yukarıya atlayın:a b Stryer, Lubert (1995). “Fotosentez”. In: Biyokimya (Dördüncü baskı). New York: WH FreeMan ve Şirketi. sayfa 653-680. ISBN 0-7167-2009-4.
- ^ Campbell, Neil A. (1990). Biyoloji (İkinci baskı). Redwood City, California: Benjamin / Cummings Publishing Company, Inc. s. 206–223. ISBN 0-8053-1800-3.
