SİNİR SİSTEMİ
| Tanımlayıcılar | |
| Latin | systema nervosum |
| MeSH | D009420 |
| FMA | 7157 |
Sinir sistemi , vücudun son derece karmaşık bir parçası olarak; canlıların kendi koordinatları doğrultusunda, işlemleri ve duyusal iletimleri, bilgi sinyalleri için ve vücudun farklı kısımlarından oluşan ileti sistemler bütününe verilen isimdir. Sinir sistemi vücudu etkileyen çevresel değişiklikleri tespit eder, daha sonra bu olaylara cevap vermek için endokrin sistemle birlikte çalışır . [1] Sinir dokusu ilk olarak yaklaşık 550 ila 600 milyon yıl önce solucan benzeri organizmalarda ortaya çıktı. Omurgalılarda merkezi sinir sistemi (CNS) ve periferik sinir sistemi olmak üzere iki ana bölümden oluşur.(PNS). CNS beyin ve omurilikten oluşur . PNS esas olarak , CNS’yi vücudun diğer bölgelerine bağlayan uzun liflerin veya aksonların kapalı demetlerinden oluşan sinirlerden oluşur . Beyinden sinyal ileten sinirlere motor veya efferent sinirler, vücuttan CNS’ye bilgi ileten sinirlere duyusal veya afferent denir. Spinal sinirler her iki fonksiyona da hizmet eder ve karışık sinirler olarak adlandırılır.
PNS üç ayrı alt sisteme ayrılmıştır: somatik , otonom ve enterik sinir sistemleri. Somatik sinirler istemli harekete aracılık eder. Otonom sinir sistemi ayrıca sempatik ve parasempatik sinir sistemlerine ayrılmıştır. Sempatik sinir sistemi enerjiyi harekete geçiren acil durumlarda aktive olurken, parasempatik sinir sistemi organizmalar rahat bir durumda olduğunda aktive olur. Enterik sinir sistemi gastrointestinal sistemi kontrol etmek için çalışır. Hem otonomik hem de enterik sinir sistemleri istemsiz çalışır. Kafatasından çıkan sinirlere kraniyal sinir, omurilikten çıkan sinirlere omurilik sinirleri denir.
Hücresel düzeyde, sinir sistemi, “sinir hücresi” olarak da bilinen nöron adı verilen özel bir hücre tipiyle tanımlanır . Nöronlar, diğer hücrelere hızlı ve hassas bir şekilde sinyal göndermelerine izin veren özel yapılara sahiptir. Bu sinyalleri, akson adı verilen ince lifler boyunca ilerleyen elektrokimyasal dalgalar şeklinde gönderirler, bu da nörotransmitter adı verilen kimyasalların sinaps adı verilen kavşaklarda salınmasına neden olur . Bir nörondan sinaptik sinyal alan bir hücre uyarılabilir, inhibe edilebilir veya başka şekilde modüle edilebilir. Nöronlar arasındaki bağlantılar sinir yolları , sinir devreleri ve daha büyük ağlar oluşturabilirbir organizmanın dünyayı algılamasını sağlayan ve onun davranışını belirleyen Sinir sistemi, nöronlarla birlikte, yapısal ve metabolik destek sağlayan glial hücreler (veya sadece glia) adı verilen diğer özel hücreleri içerir .
Sinir sistemleri çok hücreli hayvanların çoğunda bulunur, ancak karmaşıklığı büyük ölçüde değişir. [2] Sinir sistemi tek hücreli hayvanlar süngerler, placozoanlar ve mesozoanlarda çok basit bir vücut planları vardır. Sinir sistemi radyal olarak simetrik organizmalar ctenophores (tarak jöleler) ve Cnidaria üyeleri (içerir lalesi , HYDRAS , mercan ve denizanası ) diffüz oluşur. Diğer tüm hayvan türleri, birkaç solucan türü hariç olmak üzere, bir beyin, merkezi bir kablo (veya paralel olarak çalışan iki kablo ) ve beyin ve merkezi kablodan yayılan sinirleri içeren bir sinir sistemine sahiptir. Sinir sisteminin büyüklüğü en basit solucanlarda birkaç yüz hücreden Afrika fillerinde yaklaşık 300 milyar hücreye kadar değişir. [3]
Merkezi sinir sistemi bir hücreden diğerine veya vücudun bir kısmından diğerlerine sinyal göndermek ve geri bildirim almak için çalışır. Sinir sisteminin arızalanması, genetik kusurların, travma veya toksisiteye bağlı fiziksel hasar, enfeksiyon veya sadece yaşlanmanın bir sonucu olarak ortaya çıkabilir. Nörolojinin tıbbi uzmanlığı sinir sistemi bozukluklarını inceler ve bunları önleyebilecek veya tedavi edebilecek müdahaleler arar. Periferik sinir sisteminde en yaygın problem, diyabetik nöropati ve multipl skleroz ve amyotrofik lateral skleroz gibi demiyelinizan bozukluklar gibi farklı nedenlerden kaynaklanabilecek sinir iletiminin başarısızlığıdır . Nörobilim sinir sisteminin çalışılmasına odaklanan bilim alanıdır.

Omurgalı sinir sisteminin büyük bölümlerini gösteren diyagram
YAPI
Sinir sistemi adını , beyinden ve omurilikten çıkan silindirik lif demetleri ( nöronların aksonları) olan sinirlerden alır ve vücudun her parçasını innerve etmek için tekrar tekrar dallar. [4] Sinirler yeterince büyük olan eski Mısırlılar, Yunanlılar ve Romalılar tarafından tanınmış [5] ama bir mikroskop kullanarak bunları incelemek mümkün olana kadar kendi iç yapısı anlaşılamamıştır.[6] Bu konuda Yazar Michael Nikoletseas şunu yazmıştır: [7]
“Yaklaşık 1900 yılına kadar nöronların beynin temel birimleri olduğu bilinmemektedir ( Santiago Ramón y Cajal ). Aynı derecede şaşırtıcı olan şey, beyindeki kimyasal bulaşma kavramının etrafa kadar bilinmemesidir. 1930 ( Henry Hallett Dale ve Otto Loewi ). 1950’lerde nöronların kendi aralarında iletişim kurmak için kullandıkları temel elektrik fenomenini, aksiyon potansiyelini anlamaya başladık ( Alan Lloyd Hodgkin , Andrew Huxley ve John Eccles ). 1960’larda temel nöronal ağların uyaranları nasıl kodladığının ve böylece temel kavramların nasıl mümkün olduğunun farkına vardık ( David H. Hubel veTorsten Wiesel ). Moleküler devrim 1980’lerde ABD üniversitelerinde süpürüldü. 1990’larda davranışsal fenomenlerin moleküler mekanizmaları yaygın olarak biliniyordu ( Eric Richard Kandel ).”
Mikroskobik inceleme, sinirlerin öncelikle aksonlardan oluştuğunu, etraflarında saran ve fasiküllere ayrılan farklı membranlarla birlikte olduğunu gösterir. Sinirlere neden olan nöronlar tamamen sinirlerin içinde yer almazlar – hücre gövdeleri beyin, omurilik veya periferal gangliyonda bulunur . [4]
Süngerlerden daha gelişmiş olan tüm hayvanlar sinir sistemlerine sahiptir. Bununla birlikte, süngerler , tek hücreli hayvanlar ve balçık kalıplar gibi hayvanlar olmayanlar, nöronların öncüllerinden olan hücreden hücreye sinyalleşme mekanizmalarına sahiptir. [8] Denizanası ve hidra gibi radyal simetrik hayvanlarda sinir sistemi bir sinir ağından , dağınık bir izole hücre ağından oluşur [9] bu durumda bilateral var olan türlerin büyük çoğunluğunu oluşturan hayvanlar, sinir sistemi, erken kökenli ortak bir yapıya sahiptir Ediacaran 550 milyondan fazla yıl önce, dönemin. [10] [11]
HÜCRELER
Sinir sistemi iki ana kategori veya hücre tipi içerir: nöronlar ve glial hücreler.
NÖRONLAR

Dendrite; Soma; akson; çekirdek; Ranvier Düğümü; akson terminali; Schwann hücresi; Miyelin kılıf
Sinir sistemi, özel bir hücre tipi ( nöron (bazen “nöron” veya “sinir hücresi” olarak adlandırılır) varlığıyla tanımlanır . [4] Nöronlar bir dizi yolla diğer hücrelerden ayırt edilebilir, fakat bunların en temel tesis üzerinden diğer hücreler ile iletişim kurduğu bir sinaps ya sinyallerin hızla geçmesine izin veren moleküler yapıların içeren zar arası zara dezmozomlardır, elektrik veya kimyasal. [4] Birçok nöron türü , vücudun uzak bölgelerine kadar uzanabilen ve binlerce sinaptik temas kurabilen protoplazmik bir çıkıntıya sahip bir aksona sahiptir ; [12] aksonlar tipik olarak vücut boyunca sinir adı verilen demetler halinde uzanır.
İnsanlar gibi tek bir türün sinir sisteminde bile, çok çeşitli morfoloji ve işlevlere sahip yüzlerce farklı nöron türü vardır. [12] Bunlar duyu nöronları olduğu sinir sinyallerini ve de ışık ve ses gibi transmute fiziksel uyaranlar gibi motor nöron kas veya bezleri aktivasyonu gecirmislerdir sinir sinyallerin olduğu; ancak birçok türde nöronların büyük çoğunluğu merkezi yapıların (beyin ve gangliyonlar) oluşumuna katılır ve girdilerinin tamamını diğer nöronlardan alır ve çıktılarını diğer nöronlara gönderir. [4]
GLİYAL HÜCRELER
Glial hücreler (Yunancadan “tutkal” olarak adlandırılır), destek ve beslenme sağlayan, homeostazı koruyan, miyelini oluşturan ve sinir sisteminde sinyal iletimine katılan nöronal olmayan hücrelerdir. [13] de insan beyninde oranları, farklı beyin bölgelerinde farklı olsa da, glia sayısı kabaca nöron eşit olduğu tahmin edilmektedir. [14] Gliyal hücreler en önemli işlevleri arasında destek nöronlara ve bunları yerinde tutmak; nöronlara besin sağlamak; nöronları elektriksel olarak izole etmek; patojenleri yok etmekve ölü nöronları uzaklaştırmak; ve nöronların aksonlarını hedeflerine yönlendiren rehber ipuçları sağlamak. [13] , glial hücre (çok önemli bir tipi , oligodendrositlerin , merkezi sinir sisteminde ve Schwann hücreleri , periferik sinir sistemindeki) olarak adlandırılan bir yağlı madde katmanları oluşturan miyelin aksonlar etrafına sarılır ve verici aksiyon potansiyelleri izin vermesi elektrik izolasyonu sağlayan çok daha hızlı ve verimli. Son bulgular, mikroglia ve astrositler gibi glial hücrelerin, merkezi sinir sistemi içinde önemli yerleşik bağışıklık hücreleri olarak hizmet ettiğini göstermektedir.
OMURGALILARDA ANATOMİ
(CNS) ana bölümdür ve beyin ve omurilikten oluşur . [15] spinal kanal ise, omurilik içeren kafa boşluğu beyin içerir. CNS, dura mater olarak adlandırılan sert, kösele bir dış katman dahil olmak üzere üç katmanlı bir membran sistemi olan meninksler tarafından kapatılır ve korunur . Beyin ayrıca kafatası tarafından ve omurilik omurlar tarafından korunur .
Periferik sinir sistemi (PNS), CNS içinde yer almayan sinir sistemi yapıları için kolektif bir terimdir. [16] Sinir adı verilen akson demetlerinin büyük çoğunluğunun, ait oldukları nöronların hücre gövdeleri beyin veya omurilikte kalsalar bile PNS’ye ait kabul edilir. PNS somatik ve iç organlara ayrılmıştır . Somatik kısım cildi, eklemleri ve kasları sinirlendiren sinirlerden oluşur. Somatik duyu nöronlarının hücre gövdeleri dorsal kök gangliyonlarında omurilikte bulunur. Otonom sinir sistemi olarak da bilinen viseral kısım, iç organlara, kan damarlarına ve bezlere zarar veren nöronlar içerir. Otonom sinir sisteminin kendisi iki bölümden oluşur: sempatik sinir sistemi ve parasempatik sinir sistemi . Bazı yazarlar ayrıca, hücre gövdeleri PNS’nin bir parçası olarak çevre (işitme gibi duyular için) olan duyu nöronlarını; ancak diğerleri onları atlar. [17]
Omurgalı sinir sistemi gri madde ve beyaz madde olarak adlandırılan alanlara ayrılabilir . [18] Gri madde (korunmuş dokuda sadece gri olan ve canlı dokuda daha iyi pembe veya açık kahverengi olarak tanımlanan), nöronların hücre gövdelerinin büyük bir kısmını içerir. Beyaz madde esas olarak miyelinli aksonlardan oluşur ve rengini miyelinden alır. Beyaz madde tüm sinirleri ve beynin ve omuriliğin iç kısmının çoğunu içerir. Gri madde beyindeki ve omurilikteki nöron kümelerinde ve yüzeylerini hizalayan kortikal katmanlarda bulunur. Beyindeki veya omurilikteki bir nöron kümesinin çekirdek olarak adlandırıldığı anatomik bir kural vardır.buna karşılık, çevredeki nöron kümesine ganglion denir . [19] Bununla birlikte, özellikle ön beyin parçası dahil olmak üzere bu kuralın birkaç istisna olarak adlandırılan bazal ganglionlardır. [20]

Deri, kafatası ve beyni gri madde (bu görüntüde kahverengi) ve alttaki beyaz madde ile gösteren yetişkin bir dişi başın yatay bölümü
SÜNGERLERDE NÖRAL ÖNCÜLER
Süngerlerin sinaptik kavşaklarla birbirine bağlı hücreleri , yani nöronları ve dolayısıyla sinir sistemi yoktur. Bununla birlikte, sinaptik fonksiyonda anahtar rol oynayan birçok genin homologlarına sahiptirler. Son çalışmalar, sünger hücrelerinin, postsinaptik yoğunluğa (sinapsın sinyal alan kısmı) benzeyen bir yapı oluşturmak üzere birlikte kümelenen bir grup protein ifade ettiğini göstermiştir . [8] Bununla birlikte, bu yapının fonksiyonu şu anda net değildir. Sünger hücreleri sinaptik iletim göstermese de, tüm vücut kasılması gibi bazı basit eylemlere aracılık eden kalsiyum dalgaları ve diğer impulslar aracılığıyla birbirleriyle iletişim kurarlar. [21]
RADİATA
Denizanası , tarak jölesi ve ilgili hayvanlar, merkezi sinir sisteminden ziyade yaygın sinir ağlarına sahiptir. Çoğu denizanasında sinir ağı az çok vücuda eşit olarak yayılır; tarak jölelerinde ağız yakınında konsantre edilir. Sinir ağları, kimyasal, dokunsal ve görsel sinyalleri alan duyusal nöronlardan oluşur; vücut duvarının kasılmalarını aktive edebilen motor nöronlar; ve duyusal nöronlardaki aktivite modellerini tespit eden ve buna karşılık motor nöron gruplarına sinyaller gönderen ara nöronlar. Bazı durumlarda ara nöron grupları ayrı gangliyonlara kümelenir . [9]
Radiata sinir sisteminin gelişimi nispeten yapılandırılmamıştır. Bilaterianların aksine , radiata sadece iki primordial hücre tabakası vardır, endoderm ve ektoderm . Nöronlar, diğer her ektodermal hücre tipi için öncü görevi de gören özel bir ektodermal öncü hücre kümesinden üretilir. [22]
BİLATERİA

Bölünmüş genişlemelere sahip bir sinir kordonu şeklinde bir bilaterian hayvanın sinir sistemi ve ön tarafta bir “beyin”
Mevcut hayvanların büyük çoğunluğu bilaterianstır , yani sol ve sağ tarafları birbirinin yaklaşık ayna görüntüsü olan hayvanlar. Tüm bilatyaların 550-600 milyon yıl önce Ediacaran döneminde ortaya çıkan solucan benzeri ortak bir atadan geldiği düşünülmektedir. [10] Temel bilaterian vücut formu, ağızdan anüse kadar uzanan içi boş bir bağırsak boşluğuna ve her vücut segmenti için genişlemesine (“ganglion”) sahip bir sinir kordonu, özellikle beyin önde büyük bir ganglion olan bir tüptür.

İnsanlar da dahil olmak üzere memeliler bile, bölünmüş bilatiner vücut planını sinir sistemi seviyesinde gösterir. Omurilik, her biri vücut yüzeyinin bir kısmını ve altta yatan kasları sinirlendiren motor ve duyu sinirlerine yol açan bir dizi segmental gangliyon içerir. Uzuvlarda, innervasyon deseninin düzeni karmaşıktır, ancak gövdede bir dizi dar bant oluşturur. En üstteki üç segment beyine ait olup ön beyin, orta beyin ve arka beyne yol açar. [23]
Bilaterians çok erken embriyonik gelişme, iki gruba (içine meydana gelen olayların temelinde, ayrılabilir superphyla ) adı protostomes ve deuterostomes. [24] Deuterostomes omurgalıları olarak dahil derisidikenliler, yarı sırtipliler (özellikle meşe palamudu solucanlar) ve Xenoturbellidans. [25] Daha çeşitli grup olan protostomlar arasında eklembacaklılar, yumuşakçalar bulunurve sayısız solucan türü. Sinir sisteminin vücut içine yerleştirilmesinde iki grup arasında temel bir fark vardır: protostomlar vücudun ventral (genellikle alt) tarafında bir sinir kordonuna sahiptir, oysa deuterostomlarda sinir kordonu dorsal (genellikle üst) yan. Aslında, dorsal-ventral gradyanlar gösteren birkaç genin ekspresyon paternleri de dahil olmak üzere vücudun sayısız yönü iki grup arasında ters çevrilir. Çoğu anatomist şimdi protostom ve deuterostomların vücutlarının birbirlerine göre “ters çevrildiğini” düşünmektedir, bu ilk önce Geoffroy Saint-Hilaire tarafından önerilen bir hipotezdir.omurgalılara kıyasla böcekler için. Böylece böcekler, örneğin, vücudun ventral orta çizgisi boyunca uzanan sinir kordonlarına sahipken, tüm omurgalıların sırt orta çizgisi boyunca uzanan omurilikleri vardır. [26]
SOLUCAN

Solucan sinir sistemi. Üst: solucanın ön tarafının yandan görünümü. Alt: sinir sistemi izole olarak, yukarıdan izlendi
Solucanlar en basit bilaterian hayvanlardır ve bilaterian sinir sisteminin temel yapısını en basit şekilde ortaya çıkarır. Örnek olarak, solucanlarda vücudun uzunluğu boyunca uzanan ve kuyruk ve ağızda birleşen çift sinir kordonları vardır . Bu sinir kordonları, merdivenin basamakları gibi enine sinirler tarafından bağlanır . Bu enine sinirler hayvanın iki tarafını koordine etmeye yardımcı olur . Baştaki iki gangliyon (” sinir halkası “) basit bir beyine benzer şekilde işlev görür . Hayvanın gözlerindeki fotoreseptörler , aydınlık ve karanlık hakkında duyusal bilgi sağlar. [27]
Bir çok küçük yuvarlak kurt sinir sistemi, nematod Caenorhabditis elegans, tamamen içinde haritalanmıştır connectome da sinaps da dahil olmak üzere. Her nöron ve hücresel kökü kaydedildi ve sinir bağlantılarının hepsi olmasa da çoğu biliniyor. Bu türde sinir sistemi cinsel olarak dimorfiktir ; iki cinsiyetin, erkeklerin ve dişi hermafroditlerin sinir sistemleri, cinsiyete özgü işlevleri yerine getiren farklı sayıda nöron ve nöron grubuna sahiptir. Olarak C. elegans çift cinsiyetli tam olarak 302 nöronları varken, erkek, tam olarak 383 nöronlar vardır. [28]
EKLAMBACAKLILAR

Sinir sistemini mavi gösteren bir örümceğin iç anatomisi
Böcekler ve kabuklular gibi eklembacaklılar, göbek uzunluğu boyunca uzanan iki paralel bağlantıdan oluşan bir ventral sinir kordonu ile bağlanan bir dizi gangliyondan oluşan bir sinir sistemine sahiptir . [29] Tipik olarak, her vücut segmentinin her iki tarafında bir ganglion vardır, ancak bazı gangliyonlar beyin ve diğer büyük gangliyonları oluşturmak için kaynaştırılır. Baş segmenti, supraözofageal ganglion olarak da bilinen beyni içerir . Olarak böcek sinir sistemi , beyin, anatomik olarak ayrılır protocerebrum , deutocerebrum vetritocerebrum . Beynin hemen arkasında, üç çift eritilmiş gangliyondan oluşan subözofageal gangliyon bulunur. Bu kontrol ağız parçaları , tükürük bezleri ve bazı kasları . Birçok eklembacaklı, görme için bileşik gözler ve koku alma ve feromon hissi için antenler dahil olmak üzere iyi gelişmiş duyu organlarına sahiptir . Bu organlardan gelen duyusal bilgiler beyin tarafından işlenir.
Böceklerde, birçok nöronun beynin kenarına yerleştirilmiş ve elektriksel olarak pasif olan hücre gövdeleri vardır – hücre gövdeleri sadece metabolik destek sağlamaya hizmet eder ve sinyalizasyona katılmazlar. Protoplazmik bir lif hücre gövdesinden geçer ve bolca dallar, bazı kısımları sinyal iletir ve diğer kısımları sinyal alır. Böylece, böcek beyninin çoğu bölümünde, çevre çevresinde düzenlenmiş pasif hücre gövdeleri bulunurken, nöral sinyal işleme , iç kısımda nöropil adı verilen protoplazmik liflerin bir arada gerçekleşir . [30]
“TANIMLANMIŞ” NÖRONLAR
Bir nöron denir tanımlanan bu tür bir konumda, nörotransmiter, gen ekspresyonu modeli, ve aynı hayvan özelliklerinde her nöron ayırt özelliklere sahipse bağlantı-ve aynı türe ait her bir organizma bir ve sadece bir tane aynı özelliklere sahip nöron. [31] Omurgalı sinir sistemlerinde bu anlamda çok az sayıda nöron “tanımlanmıştır” – insanlarda hiçbirinin olmadığına inanılmaktadır – ancak daha basit sinir sistemlerinde, nöronların bazıları veya tümü benzersiz olabilir. Yuvarlak kurt C.’de elegansSinir sistemi herhangi bir hayvanın en iyisi olarak tarif edilen vücuttaki her nöron, her bir solucanda aynı yer ve aynı bağlantılarla benzersiz bir şekilde tanımlanabilir. Bu gerçeğin göze çarpan bir sonucu, C. elegans sinir sisteminin formunun, deneyime bağımlı bir plastisite olmadan, tamamen genom tarafından belirtilmesidir. [28]
Pek çok yumuşakça ve böceğin beyinleri de önemli sayıda tanımlanmış nöron içerir. [31] omurgalılarda, en iyi bilinen tespit nöronlar dev Mauthner hücreleri balık. [32] Her balık beyin sapı, sol tarafta biri sağda biri alt kısmında yer alan iki Mauthner hücresi var. Her Mauthner hücresi, aynı beyin seviyesinde nöronları innerve ederek omurilikten aşağı doğru ilerleyerek çok sayıda bağlantı kurarak geçen bir aksona sahiptir. Bir Mauthner hücresi tarafından üretilen sinapslar o kadar güçlüdür ki, tek bir aksiyon potansiyeli büyük bir davranış tepkisine yol açar: milisaniyeler içinde balık vücudunu C-şekline sokarsonra düzelir, böylece hızla ileriye doğru itilir. İşlevsel olarak bu, balıkların yanal çizgi organına çarpan güçlü bir ses dalgası veya basınç dalgası tarafından en kolay şekilde tetiklenen hızlı bir kaçış tepkisidir. Mauthner hücreleri balıkta tanımlanmış tek nöron değildir – her spinal segmental çekirdeğin “Mauthner hücre analogları” çiftleri de dahil olmak üzere yaklaşık 20 tip daha vardır. Her ne kadar bir Mauthner hücresi ayrı ayrı bir kaçış tepkisi getirebilse de, sıradan davranış bağlamında diğer hücre tipleri genellikle yanıtın genliğini ve yönünü şekillendirmeye katkıda bulunur.
Mauthner hücreleri komuta nöronları olarak tanımlanmıştır . Bir komut nöronu, belirli bir davranışı tek tek sürdürebilen bir nöron olarak tanımlanan özel bir tanımlanmış nöron türüdür. [33] Bu tür nöronlar en çok çeşitli türlerin hızlı kaçış sistemlerinde görülür – muazzam boyutları nedeniyle nörofizyolojide öncü deneyler için kullanılan kalamar devi aksonu ve kalamar devi sinaps , kalamarın hızlı kaçış devresine katılır. Bununla birlikte, bir komuta nöronu kavramı, başlangıçta açıklamaya uygun görünen bazı nöronların gerçekten sadece sınırlı bir dizi durumda bir cevap uyandırabildiğini gösteren çalışmalar nedeniyle tartışmalı hale gelmiştir. [34]
FONKSİYON
En temel düzeyde, sinir sisteminin işlevi bir hücreden diğerine veya vücudun bir kısmından diğerlerine sinyaller göndermektir. Bir hücrenin diğer hücrelere sinyal göndermesinin birçok yolu vardır. Birincisi, uzaktaki bölgelere yayılabilmeleri için hormon adı verilen kimyasalları iç dolaşıma serbest bırakmaktır . Bu “yayın” sinyalleme modunun aksine, sinir sistemi “noktadan noktaya” sinyaller sağlar – nöronlar aksonlarını belirli hedef alanlara yansıtır ve belirli hedef hücrelerle sinaptik bağlantılar kurar. [35] Bu nedenle, sinir sinyal hormon sinyal daha özgüllük çok daha yüksek bir seviyede yeteneğine sahiptir. Aynı zamanda çok daha hızlıdır: en hızlı sinir sinyalleri saniyede 100 metreyi aşan hızlarda hareket eder.
Daha bütünleştirici bir düzeyde, sinir sisteminin birincil işlevi vücudu kontrol etmektir. [4]Bunu, duyusal reseptörleri kullanarak ortamdan bilgi çıkararak, bu bilgiyi merkezi sinir sistemine kodlayan sinyaller göndererek, uygun bir tepki belirlemek için bilgileri işleyerek ve yanıtı etkinleştirmek için kaslara veya bezlere çıkış sinyalleri göndererek yapar. Karmaşık bir sinir sisteminin gelişimi, çeşitli hayvan türlerinin görme, karmaşık sosyal etkileşimler, organ sistemlerinin hızlı koordinasyonu ve eşzamanlı sinyallerin entegre işlenmesi gibi gelişmiş algılama yeteneklerine sahip olmasını mümkün kılmıştır. İnsanlarda sinir sisteminin karmaşıklığı dil, kavramların soyut temsili, kültür aktarımı ve insan beyninin olmadan var olmayacak insan toplumunun diğer birçok özelliğine sahip olmayı mümkün kılar.
NÖRONLAR VE SİNAPSLAR
 Sinaptik iletimde ana unsurlar. Aksiyon potansiyeli olarak adlandırılan bir elektrokimyasal dalga , bir nöronun aksonu boyunca hareket eder . Dalga sinapslara ulaştığında , hedef hücrenin zarında bulunan kimyasal reseptör moleküllerine bağlanan az miktarda nörotransmitter molekülünün salınımını tetikler .
Sinaptik iletimde ana unsurlar. Aksiyon potansiyeli olarak adlandırılan bir elektrokimyasal dalga , bir nöronun aksonu boyunca hareket eder . Dalga sinapslara ulaştığında , hedef hücrenin zarında bulunan kimyasal reseptör moleküllerine bağlanan az miktarda nörotransmitter molekülünün salınımını tetikler .
Bazı nöronlar dendrite-dendrite iletişim yeteneğine sahip olsa da çoğu nöron, aksonları yoluyla sinyal gönderir . (Aslında, amacrine hücreleri olarak adlandırılan nöron türlerinin aksonları yoktur ve sadece dendritleri aracılığıyla iletişim kurarlar.) Sinir sinyalleri, aksonlar boyunca , potansiyel olarak hücre-hücre sinyalleri üreten, potansiyel potansiyel olarak adlandırılan noktalarda hücre-hücre sinyalleri üreten bir akson boyunca yayılır. akson terminalleri yapmak sinaptik diğer hücreler ile temas. [36]
Sinapslar elektrik veya kimyasal olabilir. Elektrik sinapsları nöronlar arasında doğrudan elektriksel bağlantılar kurar, ancak [37] kimyasal sinapslar çok daha yaygın ve çok daha çeşitlidir. [38] Bir kimyasal sinaps presinaptik olarak adlandırılır sinyalleri gönderir hücresi ve sinyalleri alır hücre de postsinaptik olarak adlandırılır. Hem presinaptik hem de postsinaptik alanlar sinyal verme işlemini gerçekleştiren moleküler makinelerle doludur. Presinaptik alan, sinaptik veziküller adı verilen ve nörotransmitter kimyasalları ile dolu çok sayıda küçük küresel damar içerir . [36] Presinaptik terminal elektriksel olarak uyarıldığında, membrana gömülü bir dizi molekül aktive edilir ve veziküllerin içeriğinin, sinaptik yarık adı verilen presinaptik ve postsinaptik membranlar arasındaki dar alana salınmasına neden olur . Nörotransmiter daha sonra postsinaptik membrana gömülü reseptörlere bağlanır ve bu da aktif hale geçmelerine neden olur. [38] Reseptör tipine bağlı olarak, postsinaptik hücre üzerindeki sonuç etkisi, daha karmaşık şekillerde uyarıcı, engelleyici veya modülatör olabilir. Örneğin, nörotransmitter asetilkolinin motor nöron ile kas hücresi arasındaki sinaptik temasta salınmasıkas hücresinin hızlı kasılmasını indükler. [39] postsinaptik hücrede üzerindeki etkileri çok daha uzun olsa da, tüm sinaptik iletim süreci, bir milisaniye yalnızca bir kısmını alır (hatta belirsiz durumlarda veya bir oluşumuna sinaptik sinyal kabloları bellek iz ). [12]

Postsinaptik Yoğunluk Gerilim kapılı Ca ++ kanalı Sinaptik vezikül Nörotransmitter taşıyıcı reseptör nörotransmitter akson terminali Sinaptik yarık Dendrite
Kelimenin tam anlamıyla yüzlerce farklı sinaps türü vardır. Aslında, yüzden fazla bilinen nörotransmitter vardır ve bunların çoğunda birden fazla reseptör türü vardır. [40] Birçok sinaps birden fazla nörotransmitter kullanır; ortak bir düzenleme, glutamat veya GABA gibi hızlı etkili bir küçük moleküllü nörotransmitter ile birlikte daha yavaş etkili modülasyon rolleri oynayan bir veya daha fazla peptit nörotransmitterini kullanmaktır. Moleküler sinirbilimciler genellikle reseptörleri iki geniş gruba ayırır: kimyasal kapılı iyon kanalları ve ikinci haberci sistemleri. Kimyasal olarak kaplanmış bir iyon kanalı aktive edildiğinde, belirli tipte iyonların membran boyunca akmasına izin veren bir geçit oluşturur. İyon tipine bağlı olarak, hedef hücre üzerindeki etki uyarıcı veya engelleyici olabilir. İkinci bir haberci sistem aktive edildiğinde, hedef hücre içinde, sonuçta hücrenin uyaranlara duyarlılığını arttırmak veya azaltmak, hatta gen transkripsiyonunu değiştirmek gibi çok çeşitli karmaşık etkiler üretebilen bir dizi moleküler etkileşime başlar .
Bilinen birkaç istisnası olan Dale prensibi adı verilen bir kurala göre , bir nöron tüm sinapslarında aynı nörotransmitterleri serbest bırakır. [41] Bu does not ortalamaları olsa da, bir sinaps etkisi nörotransmitter bileşenlere bağlıdır, ancak reseptörler üzerinde aktif edilen çünkü bir nöron, tum hedeflerine aynı etki uygulaması yeterlidir. [38] Farklı hedefler farklı tipte reseptörler kullanabildiğinden (ve sıklıkla kullandıklarından), bir nöronun bir dizi hedef hücre üzerinde uyarıcı etkilere, diğerleri üzerinde engelleyici etkilere ve diğerlerinde karmaşık modülatör etkilere sahip olması mümkündür. Bununla birlikte, en yaygın kullanılan iki nörotransmitter, glutamat ve GABA, her birinin büyük ölçüde tutarlı etkileri vardır. Glutamat, yaygın olarak meydana gelen birkaç reseptör tipine sahiptir, ancak hepsi uyarıcı veya modülatördür. Benzer şekilde, GABA’nın yaygın olarak bulunan birkaç reseptör tipi vardır, ancak hepsi inhibitördür. [42] Bu tutarlılık nedeniyle glutamaterjik hücreler sıklıkla “uyarıcı nöronlar” ve GABAerjik hücreler “inhibitör nöronlar” olarak adlandırılır. Açıkçası, bu bir terminolojinin kötüye kullanılmasıdır – nöronlar değil, uyarıcı ve inhibitör olan reseptörlerdir – ancak bilimsel yayınlarda bile yaygın olarak görülür.
Sinapsların çok önemli bir alt kümesi, sinaptik güçte uzun süreli aktiviteye bağlı değişiklikler yoluyla bellek izleri oluşturabilir. [43] En iyi bilinen nöral bellek formu , NMDA reseptörü olarak bilinen özel bir reseptör türü üzerinde hareket eden nörotransmiter glutamat kullanan sinapslarda çalışan, uzun süreli güçlendirme (LTP kısaltması) adı verilen bir işlemdir . [44] NMDA reseptörünün bir “birleştirici” özelliği vardır: eğer sinapsta yer alan iki hücrenin her ikisi de yaklaşık olarak aynı zamanda aktive edilirse, kalsiyumun hedef hücreye akmasına izin veren bir kanal açılır. [45] Kalsiyum girişi, nihai olarak hedef hücredeki glutamat reseptörlerinin sayısında bir artışa yol açan, böylece sinapsın etkin kuvvetini artıran ikinci bir haberci kaskatını başlatır. Bu güç değişikliği haftalarca veya daha uzun sürebilir. 1973’te LTP’nin keşfinden bu yana, değişen koşulların neden olduğu ve değişken süreler boyunca devam eden sinaptik mukavemetteki artışları veya azalmaları içeren birçok başka sinaptik hafıza izi türü bulunmuştur. [44] ödül sistemi , güçlendirir, örneğin davranış arzu bu, bir ödül-sinyal yolunun gelen kullanımları gelen ilave bir giriş şartına bağlıdır LTP bir varyant şekli bağlıdır dopamin nörotransmitter olarak. [46] Kolektif olarak alınan tüm bu sinaptik değiştirilebilirlik biçimleri, sinirsel plastisiteye , yani sinir sisteminin ortamdaki değişikliklere uyum sağlama yeteneğine yol açar .
SİNİRSEL DEVRELER VE SİSTEMLER
Diğer hücrelere sinyal göndermenin temel nöronal fonksiyonu, nöronların birbirleriyle sinyal alışverişi yapma kabiliyetini içerir. Birbirine bağlı nöron grupları tarafından oluşturulan ağlar , özellik tespiti, örüntü oluşturma ve zamanlama da dahil olmak üzere çok çeşitli fonksiyonlara sahip olabilirler [47] ve sayısız bilgi işlemin mümkün olduğu görülmektedir. Warren McCulloch ve Walter Pitts 1943’te bir nöronun büyük ölçüde basitleştirilmiş bir matematiksel soyutlamasından oluşan yapay sinir ağlarının bile evrensel hesaplama yapabildiğini gösterdiler. [48]

Ağrı yolu İllüstrasyonu, René Descartes ‘ın Man Treatise
Bununla birlikte, elektrofizyolojinin 20. yüzyılın başlarından başlayarak ve 1940’lara kadar yüksek üretkenliğe ulaşan deneysel çalışmaları , sinir sisteminin harici bir uyaran gerektirmeden, içsel aktivite kalıpları oluşturmak için birçok mekanizma içerdiğini gösterdi. [52] nöronlar daha tam bir izolasyon, aksiyon potansiyeli veya patlamaları dizilerinin normal dizileri üretme yeteneğine sahip olduğu bulunmuştur. [53] Doğası gereği aktif nöronlar karmaşık devrelerde birbirine bağlandığında, karmaşık zamansal modeller üretme olanakları çok daha geniş hale gelir. [47] Modern bir anlayış, sinir sisteminin işlevini kısmen uyaran-cevap zincirleri açısından ve kısmen de içsel olarak üretilen aktivite modelleri açısından görür – her iki faaliyet türü de tam davranış repertuarını oluşturmak için birbirleriyle etkileşime girer. [54]
REFLEKSLER VE DİĞER UYARICI CEVAP DEVRELERİ

Temel sinir sistemi fonksiyonunun basitleştirilmiş şeması: sinyaller duyu reseptörleri tarafından alınır ve omuriliğe ve beyne gönderilir, burada işlem gerçekleşir ve bu da omuriliğe geri gönderilir ve daha sonra motor nöronlara gönderilir.
Nöral devrenin basit tip a, refleks yayı , bir ile başlar, duyusal olarak bağlı nöronların bir dizisi boyunca geçen bir motor çıkışı ile girişi ve uçları seri . [55] Bu, “geri çekme refleksinde” gösterilebilir ve sıcak soba dokunulduktan sonra bir elin geri çekilmesine neden olur. Devre duyusal reseptörlerle başlar zararlı ısı seviyeleri ile aktive edilen deride: membrana gömülü özel bir moleküler yapı tipi, ısının membran boyunca elektrik alanını değiştirmesine neden olur. Elektrik potansiyelindeki değişiklik, verilen eşiği geçecek kadar büyükse, reseptör hücresinin aksonu boyunca omuriliğe iletilen bir aksiyon potansiyelini uyandırır. Akson, bazıları omuriliğin aynı bölgesine, bazıları da beynin içine doğru çıkıntı yapan diğer hücrelerle uyarıcı sinaptik temaslar yapar. Bir hedef, bir dizi omurga interneronudurbu projeyi kol kaslarını kontrol eden motor nöronlara yönlendirdi. İnterneronlar motor nöronları uyarır ve eğer uyarma yeterince güçlü olursa, motor nöronların bazıları aksonlarını kas hücreleri ile uyarıcı sinaptik temaslar yaptıkları noktaya kadar hareket eden aksiyon potansiyelleri üretir. Uyarıcı sinyaller, kas hücrelerinin kasılmasını indükler, bu da koldaki eklem açılarının değişmesine neden olur ve kolu çeker.
Gerçekte, bu basit şema çok sayıda komplikasyona tabidir. [55] En basit refleksler için duyusal nörondan motor nörona kısa sinir yolları olsa da, devreye katılan ve yanıtı modüle eden diğer yakın nöronlar da vardır. Ayrıca, beynin omuriliğe refleksini arttırabilen veya inhibe edebilen çıkıntılar vardır.
En basit reflekslere tamamen omurilikte yatan devreler aracılık etse de, daha karmaşık tepkiler beyinde sinyal işlemeye dayanır. [56]Örneğin, görsel alanın çevresindeki bir nesne hareket ettiğinde ve bir kişi ona baktığında, sinyal işlemenin birçok aşaması başlatılır. Beyin sapının okülomotor çekirdeğindeki ilk duyusal tepki ve son retina tepkisi, basit bir refleksdekinden farklı değildir, ancak ara aşamalar tamamen farklıdır. Bir veya iki aşamalı işlem zinciri yerine, görsel sinyaller, belki de bir düzine entegrasyon aşamasından geçer ve talamus, serebral korteks, bazal gangliyonlar, üstün kolikülüs, serebellum ve birkaç beyin sapı çekirdeğini içerir. Bu alanlar, özellik algılama , algısal analiz, bellek hatırlama ,karar verme ve motor planlama . [57]
Özellik algılama , duyusal sinyallerin kombinasyonlarından biyolojik olarak ilgili bilgileri çıkarma yeteneğidir. [58] de görme sistemi , örneğin, duyu reseptörleri retina gözün sadece tek tek dış dünya “ışık noktaları” tespit etme yeteneğine sahiptir. [59] İkinci seviye görsel nöronlar birincil reseptör gruplarından girdi alır, daha yüksek seviye nöronlar ikinci seviye nöron gruplarından girdi alır ve bu şekilde işleme aşamaları hiyerarşisi oluşturur. Her aşamada, sinyal grubundan önemli bilgiler çıkarılır ve önemsiz bilgiler atılır. İşlemin sonunda, “ışık noktalarını” temsil eden giriş sinyalleri, çevredeki dünyadaki nesnelerin ve bunların özelliklerinin nöral bir temsiline dönüştürüldü. En sofistike duyusal işleme beynin içinde gerçekleşir, ancak karmaşık özellik ekstraksiyonu omurilikte ve retina gibi periferik duyu organlarında da gerçekleşir.
İÇSEL ÖRÜNTÜ OLUŞTURMA
Her ne kadar uyaran-cevap mekanizmaları anlaşılması en kolay olsa da, sinir sistemi aynı zamanda vücudu, dahili olarak oluşturulan aktivite ritimleri vasıtasıyla harici bir uyaran gerektirmeyen şekillerde kontrol edebilmektedir. Bir nöronun zarına gömülebilen voltaja duyarlı iyon kanallarının çeşitliliği nedeniyle, birçok nöron türü, izolasyonda bile, ritmik aksiyon potansiyeli dizileri veya yüksek hızda patlama ve sessizlik arasında ritmik değişimler üretebilir. . İçsel ritmik nöronlar uyarıcı veya engelleyici sinapslar ile birbirine bağlandığında, ortaya çıkan ağlar, çekici dinamikleri, periyodiklik ve hatta kaos dahil olmak üzere çok çeşitli dinamik davranışlara sahip olabilirler.. Geçici olarak yapılandırılmış bir uyarana ihtiyaç duymadan, geçici olarak yapılandırılmış çıktı üretmek için iç yapısını kullanan bir nöron ağına merkezi bir örüntü üreteci denir .
Dahili kalıp üretimi, milisaniyeden saatlere veya daha uzun bir zaman aralığında geniş bir aralıkta çalışır. Zamansal paternin en önemli türlerinden biri sirkadiyen ritmikliktir— Yani, yaklaşık 24 saatlik bir dönemle ritmik. İncelenen tüm hayvanlar, uyku-uyanıklık döngüsü gibi davranışlardaki sirkadiyen değişimleri kontrol eden nöral aktivitede sirkadiyen dalgalanmalar gösterir. 1990’lardan kalma deneysel çalışmalar, sirkadiyen ritimlerin, ekspresyon seviyesi gün boyunca yükselen ve düşen özel bir dizi genden oluşan bir “genetik saat” tarafından üretildiğini göstermiştir. Böcekler ve omurgalılar kadar çeşitli hayvanlar benzer bir genetik saat sistemini paylaşır. Sirkadiyen saat ışıktan etkilenir, ancak ışık seviyeleri sabit tutulduğunda ve başka harici güne ait ipuçları olmadığında bile çalışmaya devam eder. Saat genleri, sinir sisteminin birçok bölümünde ve birçok periferik organda ifade edilir, ancak memelilerde bu “doku saatleri” nin tümüsuprachiasmatik çekirdek .
AYNA NÖRONLARI
Bir ayna nöron , hem bir hayvan hareket ettiğinde hem de hayvan başka bir kişinin yaptığı aynı eylemi gözlemlediğinde ateş eden bir nörondur . [60] [61] [62] Böylece, nöron, sanki gözlemcinin kendisi hareket ediyormuş gibi diğerinin davranışını “yansıtır”. Bu tür nöronlar doğrudan primat türlerinde gözlenmiştir . [63] Kuşların taklit edici rezonans davranışlarına sahip oldukları gösterilmiştir ve nörolojik kanıtlar bir tür yansıtma sisteminin varlığına işaret etmektedir. [63] [64] İnsanlarda, ayna nöronlarınki ile tutarlı beyin aktivitesi, premotor kortekste , ek motor alanında ,primer somatosensoriyel korteks ve alt parietal korteks . [65] ayna sisteminin fonksiyonu çok spekülasyon bir konudur. Bilişsel sinirbilim ve bilişsel psikoloji alanındaki birçok araştırmacı, bu sistemin algılama / eylem eşleşmesi için fizyolojik mekanizma sağladığını düşünmektedir. [62] Onlar ayna nöronlar ve taklit yoluyla yeni beceriler öğrenmek için diğer insanların eylemlerini anlamak için önemli olabilir savunuyorlar. Bazı araştırmacılar ayrıca ayna sistemlerinin gözlemlenen eylemleri simüle edebileceğini ve böylece zihin teorisi becerilerine katkıda bulunabileceğini düşünüyorlar [66] [67] , diğerleri ise ayna nöronlarıdil yetenekleri. [68] Ancak, bugüne kadar, hiçbir yaygın olarak kabul nöral veya hesaplamalı modeller ayna nöron aktivitesinin böyle taklit olarak bilişsel işlevleri destekler nasıl açıklamak için ileri sürülmüştür. [69] Ayna nöronların rolü için yapılan iddiaların yeterli araştırma ile desteklenmediğine dikkat eden sinirbilimciler vardır. [70] [71]
GELİŞME
Omurgalılarda, embriyonik Merkezi sinir gelişme içerir doğum ve farklılaşmasını ait nöronlar gelen kök hücre öncülerinin, taşıma nihai konumlarına embriyo kendi yerlerinden gelen olgunlaşmamış nöronların, aşırı büyümesini aksonlar nöronlardan ve rehberlik hareketli bir büyüme konisi yoluyla postsinaptik ortakları doğru embriyo, nesil sinaps nihayet bu aksonlar ve bunların postsinaptik ortakları ve yaşam boyu arasındaki değişimler altında yatan öğrenme ve hafıza düşünülen sinapslardaki. [72]
Gelişimin erken bir evresindeki tüm bilaterian hayvanlar, bir ucu hayvan direği ve diğer ucu bitkisel kutup olarak adlandırılan polarize bir gastrula oluşturur . Gastrula, üç hücre tabakasına sahip bir disk şeklindedir, endoderm adı verilen bir iç tabaka , çoğu iç organın astarına neden olur, mezoderm adı verilen , kemiklere ve kaslara neden olan bir orta tabaka ve bir ektoderm adı verilen dış tabaka , cilde ve sinir sistemine yol açar. [73]
|
İnsan embriyo, sinir oluğu gösterilen |
İnsan embriyosundaki nöral tüpün gelişiminde dört aşama |
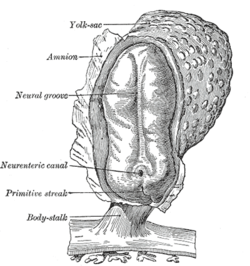

Omurgalılarda, sinir sisteminin ilk işareti, nöral plaka adı verilen sırtın merkezi boyunca ince bir hücre şeridinin ortaya çıkmasıdır . Nöral plakanın iç kısmı (orta hat boyunca), merkezi sinir sistemi (CNS), dış kısmı periferik sinir sistemi (PNS) olmaya yöneliktir. Gelişim ilerledikçe, orta hat boyunca nöral oluk adı verilen bir kat ortaya çıkar. Bu kat derinleşir ve üstte kapanır. Bu noktada gelecekteki CNS, nöral tüp adı verilen silindirik bir yapı olarak görünürken, gelecekteki PNS, nöral krest adı verilen iki doku şeridi olarak görünür, nöral tüpün üzerinde uzunlamasına çalışıyor. Sinir borusu ve nöral sinir plakadan aşamaların sırası olarak bilinen nörülasyon.
20. yüzyılın başlarında, Hans Spemann ve Hilde Mangold tarafından yapılan bir dizi ünlü deney, sinir dokusu oluşumunun organizatör bölgesi adı verilen bir mezodermal hücreden gelen sinyallerle “indüklendiğini” gösterdi . [72] Bununla birlikte, onlarca yıldır, nöral indüksiyonun doğası, bunu anlamaya yönelik her girişimi yendi, nihayet 1990’larda genetik yaklaşımlarla çözüldü. Nöral dokunun indüksiyonu, sözde kemik morfogenetik proteini veya BMP için genin inhibisyonunu gerektirir . Spesifik olarak BMP4 proteininin dahil olduğu görülmektedir. Noggin ve Chordin adlı iki proteinikisi de mezoderm tarafından salgılanır, BMP4’ü inhibe edebilir ve böylece ektodermi nöral dokuya dönüşmeye teşvik edebilir. Benzer bir moleküler mekanizmanın, eklembacaklılar ve omurgalılar da dahil olmak üzere, çok farklı hayvan türleri için rol oynadığı görülmektedir. Bununla birlikte, bazı hayvanlarda Fibroblast Büyüme Faktörü veya FGF adı verilen başka bir molekül türü de indüksiyonda önemli bir rol oynayabilir.
Nöral dokuların indüksiyonu nöroblastlar adı verilen nöral öncü hücrelerin oluşumuna neden olur . [74] Drosophila’da nöroblastlar asimetrik olarak bölünür, böylece bir ürün bir “ganglion ana hücresi” (GMC), diğeri de bir nöroblasttır. Bir GMC, bir çift nöron veya bir çift glial hücre oluşturmak için bir kez bölünür. Toplamda, bir nöroblast belirsiz sayıda nöron veya glia üretebilir.
2008 çalışmasında gösterildiği gibi, tüm ikili organizmalar (insanlar dahil) için ortak bir faktör, nörotrofinler olarak adlandırılan ve nöronların büyümesini ve hayatta kalmasını düzenleyen salgılanan sinyal molekülleri ailesidir . [75] Zhu ve diğ. sineklerde bulunan ilk nörotrofin DNT1’i tespit etti . DNT1, bilinen tüm nörotrofinlerle yapısal benzerliği paylaşır ve Drosophila’daki nöronların kaderinde anahtar bir faktördür . Nörotrofinler şimdi hem omurgalılarda hem de omurgasızlarda tanımlandığından, bu kanıtlar nörotrofinlerin iki taraflı organizmalarda ortak bir atada bulunduğunu ve sinir sistemi oluşumu için ortak bir mekanizmayı temsil edebileceğini düşündürmektedir.
PATOLOJİ

Beyin ve omuriliği koruyan katmanlar.
Her ne kadar sinirler dirsek ekleminin yakınındaki ulnar sinir gibi birkaç yer dışında derinin altında uzanma eğiliminde olsa da, hala ağrıya, his kaybına veya kas kontrolünün kaybına neden olabilecek fiziksel hasara maruz kalırlar. Sinirlere verilen hasar, karpal tünel sendromunda olduğu gibi, bir sinirin sıkı bir kemik kanalından geçtiği yerlerde şişme veya çürüklerden de kaynaklanabilir . Bir sinir tamamen kesilirse, genellikle yenilenir , ancak uzun sinirler için bu sürecin tamamlanması aylar alabilir. Fiziksel hasara ek olarak, periferik nöropati genetik durumlar, diyabet gibi metabolik durumlar da dahil olmak üzere diğer birçok tıbbi sorundan kaynaklanabilir., Guillain-Barré sendromu gibi inflamatuar durumlar , vitamin eksikliği, cüzam veya zona gibi bulaşıcı hastalıklar veya ağır metaller gibi toksinler tarafından zehirlenme. Birçok vakanın tanımlanabilecek hiçbir nedeni yoktur ve idiyopatik olarak adlandırılır . Sinirlerin geçici olarak işlevini kaybetmesi de mümkündür, bu da sertlik gibi uyuşukluğa neden olur – yaygın nedenler arasında mekanik basınç, sıcaklıkta bir düşüş veya lidokain gibi lokal anestezik ilaçlarla kimyasal etkileşimler bulunur .
Omuriliğe fiziksel hasar , duyu veya hareket kaybına neden olabilir . Omurga yaralanması şişlikten daha kötü bir şey üretmezse, semptomlar geçici olabilir, ancak omurgadaki sinir lifleri gerçekten yok edilirse, işlev kaybı genellikle kalıcıdır. Deneysel çalışmalar, omurilik sinir liflerinin sinir lifleriyle aynı şekilde yeniden büyümeye çalıştıklarını, ancak omurilikte doku yıkımının genellikle yeniden büyüyen sinirlerin nüfuz edemeyeceği skar dokusu ürettiğini göstermiştir.
KAYNAKÇA
|
