HELYCOPRİON (HELİKOPRİON)

FHPR L2003-2, bir Helicoprion davisii diş sarmalı, Phosphoria dönemine ait Idaho, Natural History Utah Field House
Helicoprion, Helcoprion veya Helikoprion olarakta bilinen, nesli tükenmiş, köpekbalığı benzeri[1] eugeneodont türü bir balıktır.
Hemen hemen tüm fosil örnekleri, yaşamda alt çeneye gömülü olan ve “diş kıvrımları” olarak adlandırılan bireylerin dişlerinin spiral olarak düzenlenmiş kümeleridir. Soyu tükenmiş kıkırdaklı balıkların çoğunda olduğu gibi, iskelet çoğunlukla bilinmemektedir. Helicoprion fosilleri 20 milyon yıl öncesiyle zamanlanmaktadır. Permiyen döneminde Artinskiyen aşamasında Cisuralian için (Erken Permiyen) Roadian aşamasında Guadalupiyende (Orta Permiyen) dönemi canlılarındandır.[2]
Helicoprion’un (ve diğer eugeneodonts) yaşayan en yakın akrabaları, ilişkileri çok uzak olmasına rağmen kimeralardır.[3] Alışılmadık diş düzeninin, yumuşak gövdeli avlarla beslenmek için bir uyarlama olduğu ve nautiloidler ve amonoidler gibi sert gövdeli kafadanbacaklılar için bir kabuk ayırma mekanizması olarak işlev gördüğü düşünülmektedir.
2013’te Helicoprion’un diş sarmallarının morfometrik analizi yoluyla sistematik revizyonu, yalnızca H. davisii, H. bessonowi ve H. ergassaminon’un geçerli olduğunu, daha büyük dişlerin bazılarının aykırı değerde olduğunu buldu. [2]
Rusya, Batı Avustralya, Çin, Kazakistan, Japonya, Laos, Norveç, Kanada, Meksika ve Amerika Birleşik Devletleri’nden (Idaho, Nevada, Wyoming, Texas, Utah ve California) bilindiği için Helicoprion’un fosilleri dünya çapında bulunmuştur.
Helicoprion’a atıfta bulunulan fosillerin %50’den fazlası, H. davisii örnekleridir. Idaho’nun Fosfor Oluşumu. Rusya’nın Ural Dağları’nda H. bessonowi türüne ait fosillerin %25’i daha bulunur. [2]
AÇIKLAMA

Helicoprion davisii’nin yeniden inşası
Diğer gibi kıkırdaklı balıklarda olduğu gibi Helicoprion ve diğer eugeneodonts iskeletler kıkırdaktan oluşmuştur. Sonuç olarak, istisnai durumlar dışında korunmamışsa, tüm vücut çürümeye başladığında parçalanmıştır. Bu, Helicoprion’un tam vücut görünümü hakkında kesin sonuçlar çıkarmayı zorlaştırmaktadır. Bununla birlikte, vücut şekli, birkaç eugeneodonts’tan bilinen postkraniyal kalıntılarla tahmin edilebilir. Korunmuş postcrania ile Eugeneodonts içerir Pensilvaniyen için Triyasik daha yukarı yaştaki caseodontoids Caseodus, Fadenia ve Romerodus bilinmektedir. [4][5] [6]
Bu taksonlar üçgen pektoral yüzgeçleri olan fusiform (aerodinamik, torpido biçimli) bir gövde planına sahiptir. Büyük ve üçgen tek vardır sırt yüzgeci, çatallı bir kanatçık omurga olmayan ve uzun bir kuyruk kanadı harici olarak görünen homocercal (iki eşit büyüklükteki loblu). Bu genel vücut planı, ton balığı, kılıç balığı ve lamnid köpekbalıkları gibi aktif, açık su yırtıcı balıklar tarafından paylaşılır.
Eugeneodonts ayrıca pelvik ve anal yüzgeçlerden yoksundur ve Romerodus’a göre, vücudun yanları boyunca kuyruk yüzgecine kadar geniş omurgaları olurdu. Fadenia muhtemelen körelmiş bir altıncı solungaç ile beş iyi açıkta solungaç yarığı vardı. Canlı kimaeroidlerde bulunan özel solungaç sepeti ve etli operkulum olduğuna dair hiçbir kanıt yoktur.[4][5] Bilinen en büyük Helicoprion diş sarmalları yaklaşık 40 cm çapındaydı. Caseodontoid diş kıvrımlarının orantılı boyutuna dayanarak, bu, Helicoprion bireylerinin modern köpekbalıklarının boyutuna rakip olarak 5 ila 8 metre uzunluğa ulaşabileceğini gösterebilir. [6]
DİŞ SARMALI
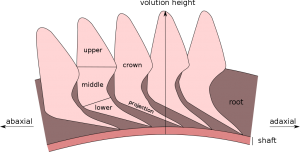
Spiral içindeki dişlerin şeması
Hemen hemen tüm Helicoprion örnekleri, yalnızca ortak bir logaritmik spiral şekilli kök içine gömülmüş düzinelerce emaye kaplı dişten oluşan “diş sarmallarından” bilinmektedir.
“Jüvenil diş kemeri” olarak adlandırılan spiralin merkezindeki en genç ve ilk diş kancalıdır, ancak diğer tüm dişler genellikle üçgen şeklindedir, yanal olarak sıkıştırılır ve sıklıkla tırtıklıdır. [7] Diş boyutu, muhtemelen 10 santimetreyi aşan en büyük dişlerle, spiralin merkezinden (aksiyal eksenden) uzaklaşır. Dişlerin alt kısmı, önceki dişin kuronunun altına çakılan çıkıntılar oluşturur. Emayeoid diş çıkıntılarının altındaki kökün en alt kısmı “şaft” olarak adlandırılır ve turbanın önceki devirlerini içine alan kıkırdak üzerinde bulunur. Tam bir diş sarmalında, spiralin en dış kısmı, diş kronunun orta ve üst kısımlarından yoksun olan uzun bir kök ile son bulur. [2]
KIKIRDAKLI KAFATASI
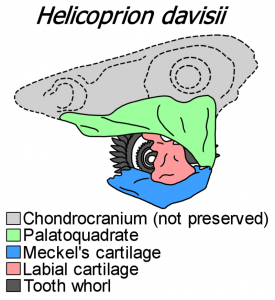
IMNH 37899’a göre Helicoprion’un kafatasının restorasyonu
Diş sarmallarından daha fazlasını koruyan Helicoprion örnekleri çok nadirdir. En iyi korunmuş numune Helicoprion (aynı zamanda “Idaho 4” olarak da bilinir) IMNH 37899 olduğu, ifade Helicoprion davisii. 1950’de Idaho’da bulundu ve ilk olarak 1966’da Svend Erik Bendix-Almgreen tarafından tanımlandı.[8]
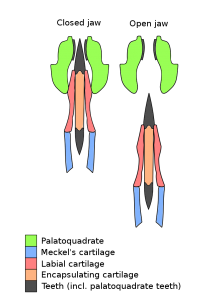
Tapanila ve meslektaşları tarafından 2013 yılında yapılan bir yeniden tanımlamaya, kıkırdaklı kalıntıları daha ayrıntılı olarak ortaya çıkarmak için BT taraması eşlik etti. BT taraması, üç boyutlu koruma ile kapalı bir konumda eklemlenmiş neredeyse eksiksiz bir çene aparatını ortaya çıkardı. Diş dövmesinin yanı sıra, örnek bir muhafaza palatoquadrate (üst çene oluşturur), Meckel kıkırdak(alt çeneyi oluşturan) ve diş kıvrımını destekleyen sağlam bir labial kıkırdak. Bu yapıların tümü, modern kondrichthyanlarda olduğu gibi prizmatik kalsifiye kıkırdaktan oluşur. Numune, beyni ve duyu organlarını barındıracak olan kıkırdaklı yapı olan bir kondrokranyumu korumadı. Çeneler, canlı kondrichthyanlara kıyasla geniş ölçüde yanal olarak (dar) sıkıştırılmıştır, ancak bu en azından kısmen ölüm sonrası sıkıştırmanın bir eseri olabilir. [3] [9]
Helicoprion’un otodiastil bir çene süspansiyonu vardı, bu da palatoquadrate’in iç kenarının kondrokranyuma iki ayrı noktada sıkıca tutturulduğu (ancak kaynaşmadığı) anlamına geliyordu. Bu iki bağlantı noktası, palatoquadrate’in önündeki kubbe şeklindeki etmoid işlem ve üst arka köşesinde flanş benzeri bazal işlemdir.[3] modern hayvanlarda sadece embriyonik bulunabilir da Autodiastylic çeneleri, ilk euchondrocephalans yaygındır chimaeriforms.[10] Bir başka iyi korunmuş numune, USNM 22577+494391 (“Sweetwood numunesi”), damak kuadratının iç yüzeyinin çok sayıda küçük (~2 mm genişliğinde) dişle kaplandığını göstermiştir.[7] Palatoquadrat dişleri alçak ve yuvarlaktı, diş kıvrımına sürtünen bir “kaldırım” oluşturuyordu. [9] Arkadan bakıldığında damak, Meckel kıkırdağı ile eşleştirilmiş bir çene eklemi oluşturur. Palatoquadrate arasındaki eklemi için hiçbir kanıt yoktur hyomandibula. [3]
Meckel’in kıkırdağı, palatoquadrat ile eklemden hemen önce ek bir çıkıntıya sahiptir. Helicoprion’a özgü bu ekstra işlem, büyük olasılıkla, çemberin kondrokranyumu delmesini önlemek için çene kapanmasını sınırlamaya hizmet etti. Helicoprion’un bir başka benzersiz özelliği, korunmuş labiyal kıkırdağın, Meckel kıkırdağının üst yüzeyi ile bir senkondroz (kaynaşmış eklem) oluşturmasıdır. Bu eklem, Meckel kıkırdağının üst kenarındaki uzun bir faset vasıtasıyla kolaylaştırılır. Labial kıkırdak, her volüsyonun köküne yakın genişleyerek diş halkası için yanal destek sağlar. Ağız kapalıyken palatoquadrata sıkışarak, labiyal kıkırdağın üst kenarı çene kapanmasının boyutunu sınırlamak için kullanılan kuvvetlerin yayılmasına yardımcı olur. Labial kıkırdağın arka kısmı, son ve en genç evrimin gelişen kökünü koruyan fincan benzeri bir forma sahiptir. [3] [9] [7]
ÖLÇEKLER
Özellikle odontodes olarak bilinen diş benzeri kondriktiyan pullarının Kazakistan’da ki H. bessonowi kalıntılarıyla ilişkili olduğu bulunmuştur. Sarcoprion ve Ornithoprion gibi diğer eugeneodontların ölçeklerine büyük ölçüde benzerler. Ölçekler, içbükey bir alt yüzeye sahip kapak şeklinde bir tabana sahiptir. Kronlar koniktir ve tırtıklı uzunlamasına sırtlarla kaplıdır. Ölçekler monodontod (her tabanda bir taç ile) veya poliodontod (birkaç odontodun daha büyük bir yapıya kaynaştırılmasından kaynaklanan çok sayıda kron demeti ile) olabilir. Diğer eugeneodonts karşılaştırıldığında, ölçekleri Helicoprion daha güçlü işaret edilmektedir. [6]
PALEOBİYOLOJİ
Helicoprion’un çenesinin kesiti . Kuyruğun iç devirleri gösterilmemiştir
Olağandışı testere benzeri diş kıvrımı ve Helicoprion’un dişlerindeki aşınmanın olmaması, sert kabuklu avın ağızdan kayacağı için yumuşak gövdeli bir av diyeti anlamına gelir. Çenenin dar yapısı nedeniyle, emme beslemesinin etkili olması pek olası değildir ve Helicoprion’un bir ısırık besleyici olduğu düşünülmektedir. Ramsay ve arkadaşları tarafından biyomekanik modelleme. (2015), sarmaldaki dişlerin spiralin neresinde olduklarına bağlı olarak farklı işlevleri olduğunu öne sürmektedir. En öndeki dişler, avı ağzına daha fazla sokmaya ve daha fazla çekmeye hizmet ederken, orta dişler mızrak ve arka dişler delinmeye ve avı boğaza daha fazla sokmaya hizmet ederken, av, turp ve palatoquadrat’ın iki yarısı arasında sıkıştı. Labial kıkırdak, turbayı desteklemeye ve desteklemeye hizmet etti. [9]
Helicoprion, ilk av yakalama sırasında büyük bir boşlukla başlamış olabilir, ardından modern ısırıkla beslenen köpekbalıkları tarafından yapıldığı gibi, avı ağza daha fazla taşımak için daha küçük çene açma ve kapama döngüleri ile başlamış olabilir. Modern köpekbalıkları, avlarını kesmeyi ve kesmeyi kolaylaştırmak için başlarını iki yana sallarken, Helicoprion’un dişleri, kesme hareketine benzer şekilde, ön dişlerin yay benzeri yolu nedeniyle, çene açılması sırasında muhtemelen avı daha da kesecektir bir bıçaktan. Helicoprion, muhtemelen, başlangıçta avı yakalamak ve ağız boşluğunun daha derinlerine itmek için bir dizi hızlı, güçlü çene kapaması kullanmış, ardından avı kesmeyi kolaylaştırmak için çenenin döngüsel açılıp kapanmasını izlemiştir. [9]

Ramsay ve meslektaşlarından sonra Helicoprion’un çene hareketi (2015)
Ramsay ve meslektaşları ayrıca, whorl’ün Erken Permiyen okyanuslarında bol miktarda bulunan amonoidler ve nautiloidler gibi sert kabuklu kafadanbacaklıların kabuklarını soymak için etkili bir mekanizma olarak hizmet edebileceğini öne sürdüler. Sert kabuklu bir kafadanbacaklı kafa kafaya ısırıldıysa, sarmaşın yumuşak gövdeyi kabuğundan çıkarıp ağzına çekmeye hizmet etmiş olması mümkündü. Çene kapanması sırasında, damak kuadratları ve diş sarmalı, üç noktalı bir sistem oluşturmak üzere birleştirilir, bu, ters çevrilmiş üç noktalı bükülme testinin kurulumuna eşdeğerdir.. Bu sistem, kesme verimliliğini artırmak ve sert kabuklu avlara karşı kaldıraç sağlamak için yumuşak parçaları yakalamada ve tutmada etkiliydi. Sahip olduğu ısırma kuvveti, çene aparatı zırhsız kemikli balıkların ve diğer chondrichthyans’ın iskelet elemanlarını kesebilecek kapasitede olduğundan, Helicoprion’un diyetini omurgalılara genişletmesine izin vermiş olabilir. [9]
SINIFLANDIRMA
IMNH 37899 den Kafatası veri yerleşimini teyit entegre hyomandibula olmadan böyle bir autodiastylic çene süspansiyon olarak çeşitli özelliklerini, ortaya Helicoprion içinde kıkırdaklı alt grup Euchondrocephalidir. Bunların aksine kardeş grubu Elasmobranchii (gerçek köpekbalığı, ışınları içeren, ve akraba), euchondrocephalans öncelikle bir sönmüş gruptur. Euchondrocephali’nin yaşayan üyeleri, yalnızca Holocephali alt sınıfındaki Chimaeriformes takımıyla temsil edilir. Yaygın olarak kimera veya sıçan balığı olarak bilinen chimaeriformlar, küçük ve özel bir nadir derin deniz kıkırdaklı balık grubudur.[11] arasındaki ilişki Helicoprion ve canlı kimeralar çok uzaktır, ancak diş anatomisinin ayrıntılarına dayanarak daha önce şüphelenilmişti. [3]
Daha spesifik olarak, Helicoprion, Devoniyen’den Triyas dönemlerine kadar yaşayan köpekbalığı benzeri bir ökondrosefalanlar takımı olan Eugeneodontida’nın bir üyesi olarak karakterize edilebilir. Eugeneodonts, düşük marjinal dişlere ve çenenin orta hattında genişlemiş bıçak benzeri simfizyal dişlere sahip basit, otodiyastil kafataslarına sahiptir. Eugeneodontida içinde, Helicoprion, özellikle uzun ve açılı simfizyal dişlere sahip bir grup eugeneodonts olan Edestoidea’ya yerleştirilir. Edestoidea üyeleri, dişlerin stiline göre iki aileye ayrılır. Bir aile, Edestidae, geriye doğru eğimli kökleri olan nispeten kısa diş bıçaklarına sahiptir.[4] [7]
İçeren diğer aile, Helicoprion, bazen cinsi dayalı Agassizodontidae denir Agassizodus.[4][7] Bununla birlikte, diğer yazarlar, ilk olarak Agassizodontidae’den 70 yıl önce kullanılan Helicoprionidae aile adını tercih ederler. Helikoprionidler (veya agassizodontidler), güçlü kemerli şekillere sahip büyük, kıkırdak destekli sarmallara sahiptir. Helikoprionidler dişlerini dökmezler; bunun yerine, diş kıvrımları, tabanları turbanın tepesinde öne doğru eğimli olacak şekilde sürekli olarak yeni dişler ekler.[4][6] Çoğu öjenodont, parçalı diş kalıntılarına dayandığından, grup içindeki somut filogenetik ilişkiler belirsizliğini koruyor. [7]
TARİH VE TÜRLER
Diş kronunun üst, orta ve alt bölümlerinin oranlarında farklılık gösteren üç tür Helicoprion morfometrik analizlerle şu anda geçerli kabul edilmektedir. Bu farklılıklar sadece spiralin 85. dişini geçen yetişkin bireylerde görülür. [2]
H. DAVİSİİ VE EŞ ANLAMLILARI

IMNH 37899 (“Idaho 4”), Phosphoria Formasyonundan bir Helicoprion davisii (= H. ferrieri ) diş sarmaşığı ve cinsteki korunmuş yumuşak dokuya sahip tek örnek
İlk örneği Helicoprion tarif edilecek olan WAMAG 9080, [2] bir kolu boyunca bulunan sarmal bir dişin 15 diş parçası Gascoyne River içinde Batı Avustralya. Henry Woodward, 1886’da fosili tanımladı ve onu keşfeden adamın anısına Edestus davisii türü olarak adlandırdı.[12] adlandırma üzerine H. bessonowi 1899, Alexander Karpinsky yeniden E. davisii için Helicoprion. [13] 1902’de Charles R. Eastman, H. davisii’ye atıfta bulundu. Yeni cinsi Campyloprion’a verildi, ancak bu teklif hiçbir zaman geniş çapta kabul görmedi.[14] Karpinsky’nin Edestus davisii’yi bir Helicoprion türü olarak tanımlaması, 1930’ların sonlarında Batı Avustralya’daki Wandagee Formasyonu’ndan birkaç daha eksiksiz diş sarmallarını tanımlayan Curt Teichert tarafından sonunda desteklenecekti. [15]
1907 ve 1909’da Oliver Perry Hay, Idaho ve Wyoming arasındaki sınırda fosfat açısından zengin Fosfor Oluşumu’nda bulunan çok sayıda fosilden yeni bir eugeneodont cinsi ve türü olan Lissoprion ferrieri’yi tanımladı. Ayrıca H. davisii’yi yeni cins ve türleriyle eş anlamlı hale getirdi. [16] Bununla birlikte, Karpinsky iki türü bir kez daha ayırdı ve 1911’de Helicoprion’a aktardı. [17] H. ferrieri başlangıçta diş açısı ve yüksekliği metrikleri kullanılarak farklılaştırıldı, ancak Tapanila ve Pruitt (2013) bu özelliklerin bu özellikleri olduğunu düşündüler. intraspesifik olarak değişkendir. Sonuç olarak yeniden atandılar H. Ferrieri bir şekilde genç eşanlamlı ait H. davisii. Fosfor Oluşumu dışında, H. davisii örnekleri Meksika, Teksas ve Kanada’da da (Nunavut ve Alberta) bulunmuştur. H. davisii, uzun ve geniş aralıklı diş kıvrımı ile karakterize edilir ve bunlar yaşla birlikte daha belirgin hale gelir. Dişler de belirgin şekilde öne doğru eğilir. [2]
1939 Yayında, Harry E. Wheeler iki yeni türler tarif Helicoprion gelen Kaliforniya ve Nevada. Bunlardan biri olan Helicoprion sierrensis, Doğu Kaliforniya’da ki buzul moren yataklarında bulunan ve muhtemelen Goodhue Formasyonu’ndan kaynaklanan bir örnekten (UNMMPC 1002) tanımlanmıştır.[18] Tapanila ve Pruitt, H. sierrensis’in ayırt edici şaft aralığının H. davisii’de bulunan varyasyon içinde olduğunu belirlediler. [2]

FMNH PF 74455, bir H. davisii (= H. Ferrieri ) diş sarmal bulunan Cam Dağları arasında Texas
H. jingmenense, 2007 yılında, bir parça ve karşı levha boyunca dörtten fazla devire sahip, neredeyse tam bir diş sarmalından (YIGM V 25147) tanımlandı. Bu Alt Permiyen aracılığıyla geçen karayolunun yapımı sırasında keşfedildi. Qixia Oluşumunda ait Hubei Eyaleti, Çin. Numune çok benzer H. Ferrieri ve H. bessonowi bu çevrim başına daha az 39 dişleri olan ikincisinden daha geniş bir kesme bıçağının, ve daha kısa bir bileşik kökü ve farklılık dişleri olan ile önceki farklı olsa da,[19] Tapanila ve Pruitt, örneğin çevredeki matris tarafından kısmen gizlendiğini ve bunun da diş yüksekliğinin olduğundan az tahmin edilmesine yol açtığını savundu. Tür içi varyasyonu hesaba katarak, bunu H. davisii ile eş anlamlı hale getirdiler. [2]
H. BESSONOWİ VE EŞ ANLAMLILARI

Rusya’nın Ural Dağları’ndan iyi korunmuş bir Helicoprion bessonowi diş sarmaşığı olan TsNIGR 1/1865 numunesinin dökümü
Helicoprion bessonowi ilk olarak Alexander Karpinsky tarafından 1899 tarihli bir monografide tanımlanmıştır. Tanımlanan ilk Helicoprion türü olmamasına rağmen, tam diş sarmallarından bilinen ilk tür olması, Helicoprion’un Edestus’tan farklı olduğunu göstermektedir. [13] bir sonucu olarak, bessonowi H. olarak hizmet türü türleri için Helicoprion.[20] H bessonowi esas alınan örnekler, bir dizi dayanır Artinskiyen daha yukarı yaştaki kireçtaşı arasında Divya formasyonu olarak, Urallar’ın arasında Rusya. H. bessonowi numuneler ayrıca bilinmektedir Tanukihara Oluşumunda ait Japonya’da[21] ve Artinskiyen çağındaki tabakalarının Kazakistan.[6] Diğer Helicoprion türlerinden, kısa ve dar aralıklı bir diş sarmalı, geriye doğru yönlendirilmiş diş uçları, geniş açılı diş tabanları ve sürekli olarak dar bir sarmal şaft ile ayırt edilebilir. [2]
Wheeler tarafından 1939’da tanımlanan iki Helicoprion türünden biri olan H. nevadensis, 1929’da Elbert A. Stuart tarafından Nevadan madeninde bulunan tek bir kısmi fosile dayanmaktadır.[18] Bu fosil, UNMMPC 1001, kaybolmuştur. Wheeler’ın Artinskian yaşında olduğunu düşündüğü Rochester Trachyte yataklarından kaynaklandığı bildirildi. Bununla birlikte, Rochester Trachyte aslında Triyas’tır ve H. nevadensis muhtemelen Rochester Trachyte’den kaynaklanmamıştır, bu nedenle gerçek yaşı bilinmemektedir. Wheeler farklılaşmış H. nevadensis gelen H. bessonowiancak Leif Tapanila ve Jesse Pruitt, 2013 yılında, örneğin temsil ettiği gelişim aşamasında bunların H. bessonowi ile tutarlı olduğunu gösterdi. [2]

H. bessonowi’nin yaşam restorasyonu
İzole diş ve kısmi kıvrımları göre adasında bulunan Spitsbergen, Norveç, H. svalis 1970 Stanislaw Siedlecki tipi numune, tarif edilen bir çok büyük bir sarmal ile PMO A-33.961, dar diş belirtilmişti örnek sayısı, bu görünüşe göre birbirleriyle temas halinde değiller. Bununla birlikte, Tapanila ve Pruitt’e göre bu, dişlerin yalnızca orta kısmının korunmasının bir sonucu gibi görünüyor. Döngü şaftı kısmen gizlendiği için, H. svalis kesin olarak H. bessonowi’ye atanamaz, ancak oranlarının birçok yönünden ikinci türe yakından yaklaşır. 72 mm maksimum devir yüksekliği ile H. svalis 76 mm maksimum devir yüksekliğine sahip en büyük H. bessonowi’ye benzer boyuttadır. [2]
H. ergassaminon

IMNH 30900 , Idaho , Bingham County’deki Gay Madeninden bir Helicoprion ergassaminon diş sarmal .
Gibi H. davisii, Helicoprion ergassaminon Idaho Phosphoria Formasyonu’ndan bilinmektedir. Ancak, nispeten çok daha nadirdir. H. ergassimon, Svend Erik Bendix-Almgreen tarafından 1966 tarihli bir monografide adlandırılmış ve ayrıntılı olarak anlatılmıştır.[8] Şimdi kayıp olan holotip numune (“Idaho 5”), beslemede kullanımının göstergesi olan kırılma ve aşınma izleri taşıyor. H. ergassimon ayrıca Fosfor Oluşumu’ndan birkaç başka örnekle temsil edilir, ancak bunların hiçbiri aşınma izi göstermez. Bu tür, H. bessonowi ve H. davisii tarafından temsil edilen iki zıt form arasında kabaca orta düzeydedir, uzun ama dar aralıklı dişlere sahip. Dişleri ayrıca geniş açılı diş tabanları ile hafifçe kavislidir. [2]
DİĞER MALZEME
H. svalis gibi belirli bir tür grubuna birkaç büyük turba atamak zordur. Idaho’dan bir örnek olan IMNH 14095, H. bessonowi’ye benziyor gibi görünüyor, ancak dişlerinin uçlarında benzersiz flanş benzeri kenarlara sahip. Yine Idaho’dan IMNH 49382, en dıştaki dönüş için 56 mm ile bilinen en büyük tur çapına sahiptir (tek korunmuş), ancak tam olarak korunmamıştır ve hala kısmen gömülüdür. [2] 1945 yılında FKG Müllerreid tarafından isimlendirilen H. mexicanus, sözde diş süslemesiyle ayırt edildi. Morfolojisi IMNH 49382’ninkine benzer olmasına rağmen, holotipi şu anda eksiktir. Başka bir materyalin yokluğunda, şu anda bir nomen dubium’dur. Vladimir Obruchev 1953’te H. karpinskii’yi iki dişinden tanımladı. Bu tür için hiçbir ayırt edici özellik sağlamadı, bu nedenle bir nomen nudum olarak kabul edilmelidir.[2] Kanada, Japonya, Laos, [22] Idaho, Utah, Wyoming ve Nevada’dan çeşitli diğer belirsiz Helicoprion örnekleri tanımlanmıştır. [20] [2]
1922 yılında Karpinsky ait yeni bir tür adında Helicoprion, H. ivanovi dan, Gzhelian (en son Karbonifer yakınında) tabakalarından Moskova. Bununla birlikte, bu tür daha sonra Helicoprion’dan çıkarılmış ve ilgili eugeneodont Campyloprion’un ikinci bir türü olarak yerleştirilmiştir. [23]
TARİHSEL REKONSTRÜKSİYONLAR
EN ERKEN REKONSTRÜKSİYONLAR
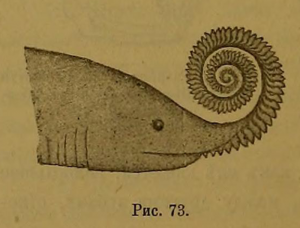
Alexander Karpinsky’nin H. bessonowi üzerindeki diş sarmalının yerleşimine ilişkin 1899 hipotezi .
Helicoprion’un diş halkalarının yerleşimi ve kimliği ile ilgili hipotezler, keşfedildiği andan itibaren tartışmalıydı. Bilinen ilk sevk Woodward(1886), Helicoprion için fosil Edestus, doğası ile ilgili hipotez ele Edestus fosil. İlk olarak Edestus vorax’ı tanımlayan Joseph Leidy, bunların “plagiostomous” (chondrichthyan) balıkların çenelerini temsil ettiğini savundu. William Davies, özellikle bir Permiyen petalodontu olan Janassa bituminosa’nın çeneleriyle karşılaştırarak kabul etti. Öte yandan, JS Newberryçene benzeri fosillerin, sicim benzeri bir balığın savunma dikenleri olduğunu öne sürdü. Woodward sonunda ED Cope’un “Pelecopterus“a (şimdi Protosphyraena olarak bilinir) benzer balıklardan elde edilen göğüs yüzgeci dikenlerini temsil ettikleri yönündeki argümanına karar verdi. [12]
Karpinsky’nin Helicoprion üzerine 1899 monografisi, diş sarmalının tuhaf doğasının işlevi hakkında kesin sonuçlara varmayı zorlaştırdığını kaydetti. Savunma veya saldırgan amaçlar için üst çeneden kıvrıldığını geçici olarak önerdi. Bu, 1899’da çeneye ait yapılar olarak yeniden değerlendirilen Edestus’un üst diş bıçaklarıyla karşılaştırılarak doğrulandı. [13]
Karpinsky’nin monografisini takip eden yıllarda Helicoprion’un diş sarmaşığının kimliğine M ilişkin tartışmalar çoktu. 1900’de yayın, makaleyi bir bütün olarak takdir eden ancak whorl’ün varsayılan yaşam pozisyonunun taslağını çıkaran Charles Eastman tarafından gözden geçirildi. Eastman, whorl’un dişlerinin diğer kondrichthyanların dişlerine çok benzediğini kabul etmesine rağmen, whorl’un ağızdan ziyade hayvanın vücuduna gömülü bir savunma yapısı olabileceği fikrini destekledi.[24] Orijinal monografisinden kısa bir süre sonra, Karpinsky, kıvrımın, Hippocampus’a (denizatı) benzeyen kıvrılmış, sivri kaplı bir kuyruğu temsil ettiği argümanını yayınladı.[25] Bu öneri, çeşitli araştırmacılar tarafından hemen eleştirildi. E. Van den Broeck yapının kırılganlığına dikkat çekti ve hayvanın yanağında çift besleme aparatı olarak en iyi şekilde korunduğunu savundu. [26] AS Woodward (Henry Woodward ile ilgisi olmayan) bu öneriyi, her bir sarmalın devasa bir köpekbalığından bir diş pilini temsil ettiği hipoteziyle takip etti. [27] G. Simoens, Karpinsky’nin çeşitli önerilerini örnekledi ve histolojik verileri, ağırşakların ağız içine yerleştirilmiş dişli yapılar olduğunu inatla savunmak için kullandı. [28] 1911’de Karpinsky, sarmalları sırt yüzgecinin bileşenleri olarak gösterdi. [17]Karpinsky’nin (1899)kine benzer yeniden yapılandırmalar, 2001’e kadar Rus yayınlarında yaygındı. [6]
DAHA SONRAKİ REKONSTRÜKSİYONLAR

H. bessonowi’nin , Janvier (1996) tarafından yayınlanan bir figüre dayanan kafatası ile modası geçmiş bir illüstrasyonu .
20. yüzyılın ortalarına gelindiğinde, diş halkası genellikle hayvanın alt çenesine yerleştirilmiş olarak kabul edildi. Karpinsky’nin monografisinden hemen sonra bu genel durumdan şüphelenilse de, 1900’lerin ortalarına kadar bu şekilde gösterilmedi. O sıralarda, yalnızca “F. John” olarak bilinen bir sanatçı, Helicoprion’u bir dizi “Tiere der Urwelt” ticaret kartı içinde tasvir etti. Onların rekonstrüksiyonu, diş kıvrımını hayvanın alt çenesinden aşağı doğru kıvrılan bir dış yapı olarak sundu. Benzer aşağı doğru kıvrılan rekonstrüksiyonlar, John A. Long, Todd Marshall ve Karen Carr gibi modern paleontologlar ve sanatçılar tarafından da yaratılmıştır.. Yeniden Bu tür diş halka dizilişli yapraklar üzerine yararı anlaşılmaktadır dayanıyordu sawfish kendi denticle kaplı snouts yanal darbeler ile incapacitate av. [6] [3]
Eugeneodont diş sarmallarının konumuyla ilgili bilgiler 1966’da iki büyük yayınla desteklendi. Birincisi Rainer Zangerl’in yeni bir Karbonifer eugeneodont, Ornithoprion tanımıydı. Bu takson, simfizyal bir pozisyonda, yani alt çenenin tabanının orta hattında küçük bir diş halkasına sahip oldukça özel bir kafatasına sahipti. O zamanlar Sarcoprion ve Fadenia için kafatası materyali de rapor edilmiş olmasına rağmen, Ornithoprion, kafatasının ayrıntılı olarak tarif edildiği ilk eugeneodont oldu. [29]
Diğer yayın üzerinde Bendix’in-Almgreen monografisi oldu Helicoprion. Araştırmaları, diş kıvrımını, öndeki bir boşlukla ayrılan meckelian kıkırdaklar arasında sıkışmış bir simfizyal yapı olarak yeniden yorumladı. Bir çift kıkırdak halkası, symphyseal crista, çene simfizinin eşleştirilmiş bir uzantısı olarak gelişiyor gibi görünüyor.meckelian kıkırdakların çenenin arkasında buluştuğu yer. Her ilmek, diş kıvrımının kökünü izleyerek içe doğru kıvrılmadan önce kavisler çizer. En büyük ve en genç dişler, çenenin arkasına yakın olan simfizde oluşur. Zamanla symphyseal crista boyunca taşınırlar, ileriye, sonra aşağıya ve içeriye doğru spiraller çizerler. Diş dizisi, simfizyal crista tarafından tanımlanan boşluk içinde yer alan spiral bir yapı içinde birikir. Diş kıvrımının yan ve alt kenarları yaşam boyunca cilt tarafından gizlenirdi. Bendix-Almgreen’e göre, diş sarmalının en olası kullanımı, yırtma ve üst çeneye karşı av kesme aracıydı. [8]
1994 tarihli Planet Ocean: A Story of Life, the Sea ve Dancing to the Fosil Record adlı kitapta, yazar Brad Matsen ve sanatçı Ray Troll, Bendix-Almgreen (1966) tarafından derlenen bilgilere dayanan bir yeniden yapılanmayı anlatıyor ve tasvir ediyor. Hayvanın üst çenesinde, sarmaşın kesmesi için dişleri kırmanın yanı sıra hiçbir diş bulunmadığını öne sürdüler. İkili, yaşayan hayvanın uzun ve çok dar bir kafatasına sahip olduğunu ve günümüz goblin köpekbalığına benzeyen uzun bir burun oluşturduğunu hayal etti.[30] Philippe Janvier’in 1996 tarihli bir ders kitabı, üst çenenin önünde keskin dişler ve çenenin arkasında sıra sıra düşük ezici dişler olmasına rağmen benzer bir rekonstrüksiyon sunmuştur. [31]
2008’de Mary Parrish, Smithsonian Doğa Tarihi Müzesi’ndeki yenilenmiş Ocean Hall için yeni bir rekonstrüksiyon yarattı. Yönetimindeki tasarlanan Robert Purdy, Victor Springer ve Matt Carrano, Parrish’in yeniden inşası, fahişeyi boğazın daha derinlerine yerleştirir. Bu hipotez, dişlerin sözde aşınma izlerine sahip olmadığı argümanı ve çemberin simfizyal bir pozisyonda yerleştirildiğinde hayvanın çenesinde sürtünmeye neden olan bir şişkinlik oluşturacağı varsayımıyla doğrulandı. Diş kıvrımını boğaz dişlerinden türetilen ve yutmaya yardımcı olmak için tasarlanmış bir yapı olarak tasavvur ettiler. Bu varsayımsal olarak, diş sarmalının çenede daha ileriye konumlandırılırsa yaratacağı dezavantajları ortadan kaldıracaktır.[32] Bu rekonstrüksiyon, yalnızca yutmaya yardımcı olmak için kullanılıyorsa, böyle bir yapının aşırı karmaşık ve potansiyel olarak etkisiz tasarımı nedeniyle eleştirildi. [33]
Lebedev (2009), Bendix-Almgreen (1966) ve Troll’ün (1994)kine benzer bir yeniden yapılandırma için daha fazla destek buldu.[6] Kazakistan’da bulunan bir diş sarmaşığı radyal çizik izlerini korudu; Whorl ayrıca varsayılan caseodontoid Campodus’unkine benzer birkaç geniş tüberküllü dişin yakınında bulundu. Lebedev’in rekonstrüksiyonu, uzun alt çenenin önünde simfizyal bir pozisyonda kıkırdak korumalı bir diş kıvrımı sundu. Ağız kapatıldığında, diş sarmalı üst çenedeki uzunlamasına derin bir cebe sığardı. Hem üst çenedeki cep hem de alt çenenin kenarları yoğun Campodus benzeri dişlerle kaplı olurdu. Durum gibi ilgili helicoprionids bildirilen Bu benzerdi Sarcoprionve Agassizodus. Gelince Helicoprion ‘ın ekolojisi, bu gibi modern deniz memelileri karşılaştırıldı Physeter (sperm balina), Kogia (cüce ve cüce sperm balinaları), Grampus (Boz Yunus) ve Ziphius (gagalı balina). Bu balık ve kalamar yiyen memeliler, genellikle alt çenenin ucuyla sınırlı olmak üzere, dişlenmeyi azaltmıştır.[6] Lebedev’in rekonstrüksiyonu, varsayımsal uzun çene ve Campodus olmasına rağmen, Helicoprion’un anatomisi hakkındaki modern görüşlere yakındır.-benzeri yanal dişlenmenin yerini CT verileri almıştır. [3]
KAYNAKÇA
|
